





Ген - структурная и функциональная единица наследственности, контролирующая развитие определённого признака или свойства. Совокупность генов родители передают потомкам во время размножения. Однако перенос генов от родителей к потомкам не является единственным способом передачи генов. В 1959 году был описан случай горизонтального переноса генов. В отличие от вертикального переноса, в горизонтальном организм передаёт гены организму, который не является его потомком. Этот способ передачи широко распространён среди одноклеточных организмов и в меньшей степени среди многоклеточных.
Гены эукариот
Отметим вначале, что у эукариотических организмов ДНК присутствует не только в ядрах, но и в органеллах - митохондриях, которые есть у всех эукариот, и хлоропластах, имеющихся у зеленых растений. По многим признакам предполагается, что орга-неллы происходят от прокариот: митохондрии от а-пурпурных бактерий, а хлоропласты - от цианобактерий. Их роднят с прокариотами многие черты белок-синтезирующего аппарата. Учитывая направленность интересов генетической инженерии, ограничимся здесь рассмотрением только ядерных генов.
Строение. Гены эукариот по строению и характеру транскрипции значительно отличаются от прокариотических генов. Их отличительной особенностью является прерывность, т. е. чередование в них последовательностей нуклеотидов, которые представлены (экзоны) или не представлены (интроны) в мРНК. Отсюда ясно, что интроны относятся к некодирующим последовательностям. Они могут располагаться не только в области, ограниченной инициирующим и терминирующим кодонами, но и вне их, в начале или в конце гена. Их длина может превышать 10 т.п.н. У низших эукариот прерывные гены составляют меньшинство всех генов (5 % у дрожжей), а у высших - большинство (94 % у млекопитающих). Отметим, что мозаичность генов найдена и в прокариотических клетках.
Эволюционно связанные гены, обладающие высокой степенью физической гомологии, образуют семейства. Белки, кодируемые такими генами, действуя одновременно или на разных этапах развития организма, выполняют одинаковые функции. Например, состав белков в а- и р-цепях гемоглобина крови млекопитающих различен у эмбриона, плода и взрослого организма, что вызвано дифференциальной экспрессией генов, входящих в а- и р-семей-ства глобиновых генов. Наряду с функционирующими генами, в семействах обнаружены нефункционирующие. Такие гены получили название псевдогенов. Они не экспрессируются по различным причинам (изменение рамки считывания из-за делеции или вставки, отсутствие интрона и т. п.).
Характерной чертой генов, входящих в семейство, является сходная картина локализации большинства интронов. Это сходство не ограничивается рамками определенного генома. Так, в случае глобиновых генов сходными по расположению интронов оказались гены у всех исследованных животных - у млекопитающих, птиц и лягушек. Однако длины и нуклеотидные последовательности интронов могут значительно варьировать, меняя тем самым и размеры самих генов.
Транскрипция. Гены эукариот не группируются в опероны, поэтому каждый из них имеет собственные промотор и терминатор транскрипции. Транскрипцию ведут три различные РНК-полимеразы: I, II и III, которые синтезируют рРНК, мРНК и тРНК, соответственно. Как и в случае прокариот, рассмотрим только механизм экспрессии генов, кодирующих белки. Поэтому далее под эукариотической РНК-полимеразой подразумевается РНК-полимераза II. Она состоит из более десятка субъединиц, но все же связываться непосредственно с промотором не может. Ее посадке на промотор способствуют транскрипционные факторы белковой природы. Ряд из них распознают специфические последовательности (боксы) в промоторе.
Длина типового промотора высших эукариот - около 100 п.н. В нем следует различать две части - базовую и дополнительную. Гены, имеющие только базовую часть промотора, функционируют в любых клетках организма и не подвержены ткане-специфичес-кому контролю. Эта часть служит для инициации транскрипции и точной ориентации РНК-полимеразы II относительно первого транскрибируемого нуклеотида. Дополнительная часть совместно с энхансерами используется для повышения эффективности транскрипции и регуляции активности гена.
Прокариоты (лат. Procaryota , от др.-греч. προ «перед» и κάρυον «ядро»), или доядерные - одноклеточные живые организмы, не обладающие (в отличие от эукариот) оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов - линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являютсяорганеллы эукариотических клеток - митохондрии и пластиды.
Прокариоты разделяют на два таксона в ранге домена (надцарства): Бактерии (Bacteria ) и Археи (Archaea ).
Для клеток прокариот характерно отсутствие ядерной оболочки, ДНК упакована без участия гистонов. Тип питания осмотрофный.
Генетический материал прокариот представлен одной молекулой ДНК, замкнутой в кольцо, имеется только один репликон. В клетках отсутствуют органоиды, имеющие мембранное строение. В геноме могут присутствовать мобильные генетические элементы, а у некоторых прокариот (например, вольбахия) их содержится необычно много. Изучение бактерий привело к открытиюгоризонтального переноса генов, который был описан в Японии в 1959 г. Это процесс широко распространен среди прокариот, а также у некоторых эукариот. Открытие горизонтального переноса генов у прокариот заставило по-другому взглянуть на эволюцию жизни. Ранее эволюционная теория базировалась на том, что виды не могут обмениваться наследственной информацией. Прокариоты могут обмениваться генами между собой непосредственно (конъюгация, трансформация) а также с помощью вирусов -бактериофагов (трансдукция).
Уникальные гены - это гены, которые встречаются в клетке два или несколько раз (до 10-20). Большинство исследователей считает, что у многоклеточных общее число генов в среднем равно сто тысяч и подавляющее их число - это уникальные гены. Характерная черта генов эукариотов - мозаичное экзон-интронное строение. Интроны, не несущие генетической информации, вырезаются (сплайсинг). Число и размер интронов у разных видов варьируется. Присутствие их в гене приводит к значительному увеличению размеров гена. Интроны стабилизируют экзоны, однако существует представление, что интрон - это так называемая «эгоистическая» ДНК, не дающая организму никаких эволюционных преимуществ. Экзоны контролируют синтез белков: 1 экзон - 1 домен .
К повторяющимся генам относятся прежде всего гены больших и малых рРНК и гистонов. Число их сильно варьирует и может достигать более 2000. Гены больших рРНК организованы в блоки, в которых последовательно идут гены 18S рРНК, 58S рРНК и 28S рРНК. Между ними имеются промежутки, различающиеся по длине у разных организмов. Межгенные участки имеют повторы разных типов, с необычной последовательностью, богатых парами ГЦ. Гены низкомолекулярных ядерных РНК блоков не образуют. Гены гистонов повторяются в геноме десятки (у млекопитающих), и сотни (у дрозофилы), и тысячи (у аксолотля) раз. Причем не удается уловить связи между этим показателем и положением организма на эволюционной лестнице.
Перестраивающиеся, или рекомбинирующие, гены - это гены, кодирующие легкие и тяжелые цепи белков иммуноглобулинов , выполняющих функции антител. Гены этих белков состоят из двух типов генов для легких и пяти типов - для тяжелых цепей. Легкие цепи кодируются тремя отдельными генетическими элементами, тяжелые - четырьмя. Перестройки генома приводят к соединению разных участков и в итоге - к образованию иммуноглобулинов разных классов.
Прыгающие гены, или транспозоны, - мобильные генетические элементы . Являясь нормальным компонентом генома, они составляют его значительную часть (у дрозофилы- 7% генома), могут быть представлены многими копиями, рассеянными по геному, и имеют варьирующую локализацию. Структура разных классов мигрирующих элементов (МЭ) варьирует, но для всех их характерно наличие на концах обращенных повторов. В середине МЭ могут иметь уникальные последовательности. МЭ проявляют высокую локусную специфичность, так как могут встраиваться в определенную последовательность на хромосоме.
Повторы на днк
Повторяющаяся ДНК - атрибут любой генетической системы. Еще на самых ранних этапах эволюции, когда возник почти полный репертуар полипептидов - носителей основных молекулярных функций (что-то вроде словаря генетического языка), им уже, по всей видимости была присуща внутренняя периодичность ( Ohno, 1981 , 1984 ; Go, 1983 ). Наличие олигомерных повторов в современных генах и белках объясняют тем, что вообще в любой информационной системе копирование текста - эффективное средство повышения помехоустойчивости при передаче сообщений . Применительно к генетическому языку существование внутригенных повторов резко снижает негативные эффекты таких ошибок, как сдвиги рамки трансляции, различные делеции и вставки ( Ohno, 1984 ). Действительно, белки, чьи гены содержат внутренние олигомерные повторы, также должны обладать периодичной первичной структурой. При этом длина полипептидного повтора будет зависеть от того, кратна ли трем длина соответствующего матричного повтора. Например, повторы из шести нуклеотидов будут порождать исключительно дипептидную периодичность, тогда как пентамерным повторам в гене должна отвечать периодичность той же размерности и в полипептидном продукте ( рис 1 ).
Но, пожалуй, самая интересная особенность таких повторов состоит в том, что они обеспечивают совпадение обеих периодичностей, нуклеотидной и аминокислотной, во всех трех возможных рамках трансляции ( Ohno, 1984 , 1987 ). Поэтому С.Оно и полагает, что еще в самом начале, на заре жизни, когда биохимическая машина трансляции работала с частыми сбоями и когда рамка считывания вряд ли была однозначно фиксирована, как раз такие повторы (длиной, не кратной трем) должны были иметь ощутимое селективное преимущество. В изрядном количестве олигонуклеотидные мотивы именно такой конфигурации присутствуют практически во всех проанализированных генах самых разных видов про- и эукариот. В принципе нельзя исключить, что по крайней мере некоторые из них сохранились с тех давних времен, т.е. являются своеобразными молекулярными реликтами.
Далее, спонтанно возникнув, даже сравнительно короткие повторенные участки существенно увеличивают вероятность дупликации (и автоматически - мультипликации) как их самих, так и тех сегментов генома, которые ими фланкированы ( Smith, 1976 ). Конкретные механизмы внутригеномного размножения повторов могут быть разными (неравный кроссинговер, скользящая гиперрепликация, обратная транскрипция и т.д.), но само размножение представляет собой типичный автокаталитический процесс ( Orgel, Crick, 1980 ; Doolitle et al., 1984 ; и др.). И хотя в целом ряде случаев количество повторов (например, МГЭ) регулируется по принципу обратной связи ( O"hare, Rubin, 1983 ; Simons, Kleckner, 1983 ; Snyder, Doolitle, 1988 ), хотя для любого вида организмов в норме имеются, по-видимому, и неспецифические селективные барьеры на пути "эгоистичного" распространения повторов по геному ( Бердников, Родин, Жарких, 1982 ; Rodin et al., 1985 ; Родин, 1985а,б ), для большинства современных организмов, в особенности эукариотических, характерна чрезвычайно высокая концентрация повторов практически на всех уровнях молекулярно-генетической организации.
Структурные гены - уникальные компоненты генома, представляющие единственную последовательность, кодирующую определённый белок или некоторые виды РН……..
Вся последовательность нуклеотидов гена принимает участие в кодировании последовательности иРНК, однако часть участков иРНК в процессе ее так называемого созревания вырезают специальные ферменты, а те, что остались, образуют вторичную иРНК, из матрицы которой, собственно, и происходит синтез белков.
Участки ДНК, которые в процессе созревания иРНК вырезаются, получили название интронов , а те, что потом сшиваются и служат матрицей для трансляции, - экзонов . Как правило один ген содержит от трех до пяти интронов. Однако следует отметить, что в генах прокариот интронов и экзонов нет.
Наукой пока не известна причина сложного строения генов прокариот. Однако есть несколько факторов объясняющих это. Поскольку интронная - экзонных организация является свойством генов только эукариот, вполне обоснованное предположение: такая сложная структура генов - прогрессивное эволюционное приспособление эукариотических организмов. Считают, что, прежде всего, это может быть механизмом, который ограничивает мутационный процесс. При этом интроны выполняют функцию «ловушек» мутаций. Ведь изменение нуклеотидных последовательностей в частях структурного гена, не кодируется, не приведет к мутациям и появлению аминокислотных замен. Кроме того, если в одном из экзонов и произойдет вставка или выпадение нуклеотида, то это повлечет смещение рамки синтеза не всей иРНК, а только какой-то ее части, то есть эффект будет не таким уж и губительным. Очевидно, сложная структура гена обеспечивает его более высокую стабильность и надежность функционирования.
Кроме структурных генов, кодирующих ту или иную форму РНК, в геноме всех организмов есть еще и регуляторные гены, которые определяют начало, скорость и последовательность процессов синтеза РНК на матрице ДНК. Они являются местом прикрепления ферментов и других белков, участвующих в репликации и транскрипции, регулируют активность генов. Регуляторные гены небольшие, включают только 20-80 пар нуклеотидов каждый, а поэтому сравнительно со структурными генами занимают гораздо меньше места в геноме. Однако без этих генов, которые не кодируют специфические белки, а только регулируют процессы репликации ДНК, взаимодействие ДНК с определенными белками и ферментами, прохождения конъюгации хромосом, так же невозможно функционирование генетического аппарата, как без гормонов (веществ, синтезированных организмом в минимальных количествах) - жизнедеятельность человеческого организма.
- Промотор - регуляторная последовательность ДНК (не транскрибируется), расположенная слева от точки начала транскрипции.
- 5’ – нетранслируемая область (лидер) – начинается от точки начала транскрипции до старт-кодона (транскрибируется, но не транслируется, входит в состав зрелой мРНК)
- Кодирующая область - экзоны и интроны (транскрибируется, но интроны вырезаются из пре-мРНК, в зрелой РНК остаются только экзоны)
- 3’ – нетранслируемая область (трейлер) – начинается от стоп кодона до последовательности АААУАА (участок полиаденилирования).
Экзоны и интроны . Большинство генов эукариот имеет прерывистое строение, они содержат кодирующие последовательности – экзоны, и некодирующие последовательности - интроны.
Гены начинаются и заканчиваются экзонами, набор интронов может быть любой. Например, глобиновые гены, имеют 3 экзона и 2 интрона. Другие гены могут иметь большое количество интронов (20 и более). Например, ген, кодирующий синтез белка-рецептора для липопротеинов низкой плотности (LDL), имеет 40 интронов, а ген белка дистрофина – 79 интронов. Размеры интронов больше, чем размеры экзонов, поэтому экзоны составляют очень небольшую часть.
В зрелой молекуле мРНК присутствуют только экзоны, а интроны вырезаются из первичного транскрипта (пре-РНК) в процессе сплайсинга . Поэтому размеры зрелых мРНК эукариот всегда меньше размеров самого гена и пре- РНК.
Не все гены эукариот содержат интроны . Не содержат интронов гены гистоновых белков, гены интерферонов, гены митохондрий млекопитающих и человека.
Экзоны – это кодирующие последовательности ДНК генов эукариот, представленные в зрелой молекуле РНК.
Интроны – это некодирующие участки генов эукариот, которые транскрибируются, но затем вырезаются из первичного транскрипта во время сплайсинга и не входят в состав зрелых РНК, т.е. не транслируются.
Новая концепция гена . Благодаря альтернативному сплайсингу из одной молекулы первичного транскрипта (пре-мРНК) можно получить несколько различных вариантов молекул зрелых мРНК с разным набором экзонов. В связи с этим можно говорить о новой концепции гена: один ген – много РНК – много полипептидов и сформулировать новое определение гена. Ген - это участок молекулы ДНК, кодирующий синтез одной или нескольких функциональных молекул РНК.
Определение тонкой структуры гена, т.е. его организации, а также принципы работы, т.е. регуляции активности (включение-выключение), первоначально были установлены для прокариотических клеток.
Эти работы выполнили Франсуа Жакоб и Жак Моно (1961; Нобелевская премия 1965). Согласно концепции Жакоба–Моно, единицей регуляции активности генов у прокариот является оперон. Оперон — функциональная единица генома у прокариот, в состав которой входят цистроны (единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами, т.е. размеры оперона превышают размеры кодирующих последовательностей ДНК. Такая функциональная организация позволяет эффективнее регулировать экспрессию (проявление) этих генов.
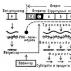
В целом структура оперона включает: промотор, оператор, структурные гены, терминатор (рис.1).

П — Промотор – это регуляторный участок ДНК, который служит для присоединения РНК-полимеразы к молекуле ДНК.
С- Оператор – это регуляторный участок ДНК, который способен присоединять белок-репрессор, который кодируется соответствующим геном. Если репрессор присоединен к оператору, то РНК-полимераза не может двигаться вдоль молекулы ДНК и синтезировать мРНК.
Т- Терминатор – это регуляторный участок ДНК, который служит для отсоединения РНК-полимеразы после окончания синтеза мРНК.
Транскрипция группы структурных генов, регулируется двумя элементами – геном-регулятором и оператором. Оператор часто локализуется между промотором и структурными генами; ген-регулятор может локализоваться рядом с опероном или на некотором расстоянии от него.
Если продуктом гена-регулятора является белок-репрессор, его присоединение к оператору блокирует транскрипцию структурных генов, препятствуя присоединению РНК-полимеразы к специфичному участку – промотору, необходимому для инициации транскрипции. Напротив, если белком-регулятором служит активный апоиндуктор, его присоединение к оператору создает условия для инициации транскрипции. В регуляции работы оперонов участвуют также низкомолекулярные вещества – эффекторы, выступающие как индукторы либо корепрессоры структурных генов, входящих в состав оперонов.
Опероны по количеству цистронов делят на моно-, олиго- и полицистронные, содержащие, соответственно, только один, несколько или много цистронов (генов).
Объединение функционально близких генов в опероны, видимо, постепенно сложилось в эволюции бактерий по той причине, что у них перенос генетической информации обычно осуществляется небольшими порциями (например, при трансдукции или посредством плазмид). Значение имеет само по себе сцепление функционально родственных генов, что позволяет бактериям приобретать необходимую функцию в один этап.
Ген - структурная и функциональная единица наследственности живых организмов. Ген представляет собой последовательность ДНК, задающую последовательность определённого полипептида либо функциональной РНК. Гены (точнее, аллели генов) определяют наследственные признаки организмов, передающиеся от родителей потомству при размножении. При этом некоторые органеллы (митохондрии, пластиды) имеют собственную ДНК, не входящую в геном организма, которая определяет их признаки.
Среди некоторых организмов, в основном одноклеточных, встречается горизонтальный перенос генов, не связанный с размножением.
Термин «ген» был введён в употребление в 1909 году датским ботаником Вильгельмом Йогансеном.
Изучением генов занимается наука генетика, родоначальником которой считается Грегор Мендель, который в 1865 году опубликовал результаты своих исследований о передаче по наследству признаков при скрещивании гороха. Сформулированные им закономерности впоследствии назвали Законами Менделя.
Среди учёных нет единого мнения под каким углом рассматривать ген. Одни учёные его рассматривают как информационную наследственную единицу, а единицей естественного отбора является вид, группа, популяция или отдельный индивид. Другие учёные, как например Ричард Докинз в своей книге «Эгоистичный ген», рассматривают ген как единицу естественного отбора, а сам организм - как машину для выживания генов.
В настоящее время, в молекулярной биологии установлено, что гены - это участки ДНК, несущие какую-либо целостную информацию - о строении одной молекулыбелка или одной молекулы РНК. Эти и другие функциональные молекулы определяют развитие, рост и функционирование организма.
В то же время, каждый ген характеризуется рядом специфических регуляторных последовательностей ДНК (англ.)русск., таких как промоторы, которые принимают непосредственное участие в регулировании проявления гена. Регуляторные последовательности могут находиться как в непосредственной близости от открытой рамки считывания, кодирующей белок, или начала последовательности РНК, как в случае с промоторами (так называемые cis cis-regulatoryelements ), так и на расстоянии многих миллионов пар оснований (нуклеотидов), как в случае с энхансерами, инсуляторами и супрессорами (иногда классифицируемые как trans -регуляторные элементы, англ. trans-regulatoryelements ). Таким образом, понятие гена не ограничено только кодирующим участком ДНК, а представляет собой более широкую концепцию, включающую в себя и регуляторные последовательности.
Изначально термин ген появился как теоретическая единица передачи дискретной наследственной информации. История биологии помнит споры о том, какие молекулы могут являться носителями наследственной информации. Большинство исследователей считали, что такими носителями могут быть только белки, так как их строение (20 аминокислот) позволяет создать больше вариантов, чем строение ДНК, которое составлено всего из четырёх видов нуклеотидов. Позже было экспериментально доказано, что именно ДНК включает в себя наследственную информацию, что было выражено в виде центральной догмы молекулярной биологии.
Гены могут подвергаться мутациям - случайным или целенаправленным изменениям последовательности нуклеотидов в цепи ДНК. Мутации могут приводить к изменению последовательности, а следовательно изменению биологических характеристик белка или РНК, которые, в свою очередь, могут иметь результатом общее или локальное изменённое или анормальное функционирование организма. Такие мутации в ряде случаев являются патогенными, так как их результатом является заболевание, или летальными на эмбриональном уровне. Однако, далеко не все изменения последовательности нуклеотидов приводят к изменению структуры белка (благодаря эффекту вырожденности генетического кода) или к существенному изменению последовательности и не являются патогенными. В частности, геном человека характеризуется однонуклеотидными полиморфизмами и вариациями числа копий (англ. copynumbervariations ), такими как делеции и дупликации, которые составляют около 1 % всей нуклеотидной последовательности человека. Однонуклеотидные полиморфизмы, в частности, определяют различные аллели одного гена.
Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие в себя азотистые основания: аденин(А) или тимин(Т) или цитозин(Ц) или гуанин(Г), пятиатомный сахар-пентозу-дезоксирибозу, по имени которой и получила название сама ДНК, а также остаток фосфорной кислоты. Эти соединения носят название нуклеотидов.
Гены и мемы
По аналогии с генами Ричардом Докинзом был введён в употребление термин «мем» - единица культурной информации. Если ген распространяется в химической среде, используя для размножения химические вещества, то мем распространяется в информационной среде: на носителях информации, в человеческой памяти, а также в сети. Также как гены конкурируют между собой за ресурсы: химические вещества, так и мемы конкурируют за информационное пространство. По целому ряду причин, между пространственным распределением генов и мемов могут наблюдаться достаточно жёсткие корреляции.
Свойства гена
2. дискретность - несмешиваемость генов;
3. стабильность - способность сохранять структуру;
4. лабильность - способность многократно мутировать;
5. множественный аллелизм - многие гены существуют в популяции во множестве молекулярных форм;
6. аллельность - в генотипе диплоидных организмов только две формы гена;
7. специфичность - каждый ген кодирует свой признак;
8. плейотропия - множественный эффект гена;
9. экспрессивность - степень выраженности гена в признаке;
10. пенетрантность - частота проявления гена в фенотипе;
11. амплификация - увеличение количества копий гена.
Классификация
В зависимости от выполняемых функций гены делятся на
1.Структурные гены – гены, контролирующие синтез структурных белков или ферментов
2. Регуляторные гены – гены, контролирующие синтез различных белков, влияющих на активность структурных генов. Регуляторные гены в свою очередь делятся на:
Гены – модификаторы – усиливающие и снижающие активность структурных генов.
Гены – супрессоры – подавляющие активность структурных генов
По влиянию на жизнеспособность организмов гены делятся на:
1 Летальные гены – гены, приводящие к гибели их носителей
Вопрос 71.Структура гена у прокариот. Оперон.
Сублитальные гены – гены, приводящие к нарушению репродуктивной функции (стерильность, пониженная жизнеспособность или нежизнеспособность потомства) их носителей
3. Нейтральные гены – не влияющие на жизнеспособность организма.
Строение структурных генов прокариот и эукариот специфичное. У прокариот в большинстве случаев кодирующий участок непрерывен, в генах эукариот наряду с участками, кодирующими специфический для этого гена продукт (полипептид, рибосомную РНК, транспортную РНК), имеются некодирующие участки. Кодирующие участки гена получили, как уже упоминалось, название экзонов, некодирующие - интронов. В структурном гене экзоны чередуются с интронами. Ген как бы разорван.
Число и внутригенная локализация интронов характерны для каждого гена. Размеры интронов различные (от нескольких десятков до нескольких тысяч нуклеотидных пар). Нередко на долю интронов в гене приходится больше нуклеотидов, чем на долю экзонов. Роль интронов мало изучена. Если бы они не выполняли определенных функций, были не нужны организму, элиминировались бы естественным отбором.
Изучение гена продолжается. Современные сведения позволяют говорить о гене как об участке молекулы геномной нуклеиновой кислоты, представляющем единицу функции и способном изменяться и приобретать различные состояния путем мутирования и рекомбинаций. Это сложная, но в функциональном отношении целостная единица наследственности.
Эукариотические гены, в отличие от бактериальных, имеют прерывистое мозаичное строение. Кодирующие последовательности (экзоны) перемежаются с некодирующими (интронами).
Генетический аппарат клеток эукариот.
Генетический аппарат клетки.
- геном – генетический материал ядра в гаплоидном наборе хромосом;
Функциональная единица – ген.
- Плазмон – генетический материал цитоплазмы;
Функциональная единица – плазмоген.
1962 год – Д. Гердон – лягушки, опыт, стоял у истоков клонирования животных.
Роль хромосом в наследственности:
- 1882 – Флемин описал поведение хромосом во время митоза;
- 1902 – Теодор Бовери, Вальтер Сеттон предположили, что гены находятся в хромосомах;
- 1909 – Томас Морган, Карл Бриджес, Альфред Стертевант экспериментально доказали связь наследственного материала с хромосомами.
Хромосомная теория наследственности.
1. Каждая хромосома представляет уникальную группу сцепления генов. Число групп сцепления равно гаплоидному набору хромосом.
2. Гены в хромосоме располагаются в линейном порядке и занимают определенное место – локус.
3. М-ду гомологичными хромосомами возможен обмен аллельными генами – кроссинговер , который нарушает сцепление генов и обеспечивает перекомбинацию генов
4. Частота кроссинговера является функцией расстояния между генами:
чем больше расстояние между генами, тем больше вероятность кроссинговера.
5. Частота кроссинговера зависит от силы сцепления м-ду генами:
чем сильнее сцеплены гены, тем меньше вероятность кроссинговера (полное и неполное сцепление).
Карл Эрих Корренс (1908) – опыты с «ночной красавицей», у которой описано явление пестролистности. Неравномерность окраски листьев объясняется неравномерным распределением хлоропластов во время деления.
Борис Эфрусси открыл метохондриальную наследственность у млекопитающих в 1949.
В 1981 была секвенирована митахондриальная ДНК человека (определена точная нуклеотидная последовательность)
ДНК митохондрий.
- Кольцевая двуспиральная;
- Содержит 37 генов:
кодируют 13 белков, 22 молекулы т-РНК, 2 молекулы р-РНК;
- Гены не содержат интронов;
- Признаки наследуются по материнской линии и не являются менделирующими.
Объем митохондриального генома в 200 тысяч раз меньше ядерного.
- Реплицируется независимо от ядерной ДНК.
- Постоянство присутствия в клетке,
- способность к самоудвоению,
- равномерное распределение генетической информации м-ду дочерними клетками во время деления.
Химический состав хромосом.
- Гистоновые белки – Н1, Н2а, Н2в, Н3, Н4 обладают основными свойствами
- Негистоновые белки обладают кислыми свойствами
- Липиды (фосфолипиды, свободные жк, хс и тг)
- Полисахариды
- Ионы металлов
Деспирализованная форма существования хромосом в неделящемся ядре называется хроматина.
Дезоксирибонуклепротеиновый комплекс (ДНП).
Степень компактизации хроматина изменяется в течении метотического цикла клетки и определяет генетическую активность или неактивность хромосом.
Чем выше степень компактизации, тем меньше генетическая активность.
Уровни компактизации:
1. Нуклеосомный:
Может быть получен только искусственным путем.
Хроматиновая фибрилла выглядит в виде ниточки бус.
Гистоновые белки 4х классов (H2a, H2b, H3 и Н4) образуют гистоновые актамеры.
На гистоновые актамеры накручивается молекула ДНК, делая 1,75 оборота.
Есть свободный линкерный участок.
В таком состоянии молекула ДНК укорачивается в6-7 раз.
Диаметр фибриллы 10 нм.
Характерен дляG1 периода интерфазы.
2. Нуклеомерный .
Хроматиновая фибрилла приобретает структуру соленоида за счет соединения соседних нуклеусов за счет встраивания белка Н1 в линкерную область.
Диаметр фибриллы 30 нм.
Коэффициент компактизации – 40 раз.
Характерен для G2 периода интерфазы.
3. Хромомерный.
Компактизация происходит при участии негистоновых белков с образованием петель.
2. Понятие о гене. Структурная организация генов прокариот и эукариот. Классификация генов.
Характерен для начала профазы митоза.
Диаметр фибриллы 300 нм.
Коэффициент компактизации 200-400 раз.
4. Хромонемный .
Петли укладываются в стопки.
Коэффициент компактизации – 1000 раз.
Диаметр фибриллы 700 нм.
Характерен для конца профазы митоза.
5. Хромосомный.
Достигается максимальная степень спирализации хроматина.
диаметр фибриллы 1400 нм.
Коэффициент компактизации 104-105.
Характерен для метафазы митоза.
Строение метафазной хромосомы.
Состоит из 2х хроматид, соединенных первичной перетяжкой или центромерой. В области центромеры находится кинетохор – участок, к которому прикрепляются нити веретена деления. Первичная перетяжка определяет форму хромосомы, деля её на 2 плеча: р – короткое плечо, q – длинное.
По форме:
- Метацентрические p=q
- Субметацентрические p
- Акроцентрические p<
- Акроцентрические p<
Для точной идентификации хромосом используют центромерный индекс: отношение длины короткого плеча к длине всей хромосомы.
Ещё есть спутники (в акроцентрических у человека), соединенные вторичной перетяжкой.
Во вторичной перетяжке находятся гены, отвечающие за синтез рибосомальных рнк.
Теломеры – концевые участки хромосом.
Роль теломер:
- Механическая функция (прикрепляются к оболочке ядра), предотвращают слипание хромосом м-ду собой, что может привести к образованию дицентриков.
- Стабилизационная – защищает хромосомы от деградации клеточными нуклеазами (ферменты, которые разрушают)
- Влияют на экспрессию генов – активность генов, расположенных рядом с теломерами снижена
- Регулируют кол-во клеточных делений в отсутствие теломеразы.
Разновидности хроматина:
1. Эухроматин — слабо компактизованные, генетически активен, реплицируется в начале интерфазы, преобладают аденин, тимин, содержит все структурные гены
1 и 2 уровень компактизации
2. Гетерохроматин – сильно компактизованные, генетически неактивен, реплицируется позже хроматина, гуанин цитозин преобладают, входят другие классы генов. Потеря участков не влияет.
3 и 4 уровень компактизации.
- Постоянный: расположен в теломерных участках хромосом и расположен в области центромеры, функции: регулирует работу структурных генов, участвует в образовании синаптического комплекса м-ду гомологичными хромосомами во время мейоза.
- Факультативный: временно переведенный в неактивное состояние эухроматин, примером его является половой хроматин.
Половой хроматин.
1949 М. Барр и Л. Бертрам обнаружили половой хроматин в интерфазных ядрах нейронов кошек.
В норме число глыбок полового хроматина на 1 меньше чем число Х-половых хромосом.
Женская зигота млекопитающих имеет две функционально активные Х-хромосомы
На 16й день эмбриогенеза происходит инактивация одной х-хромосомы во всех соматических клетках эмбриона. Процесс инактивации носит случайный характер.
Значение полового хроматина.
Тест, Только в соматических клетках! Для определения пола плода! Для диагностики хромосомных заболеваний, связанных с изменением числа половых хромосом.
В большом спорте как секс-контроль.
Кариотип – хромосомный комплекс соматических клеток определенного вида растений и животных
Показатели:
- число,
- формы и
- размеры от 0,1 до 10 мкм хромосом.
1956 – Ю. Тио и А. Леван изучили кариотип человека.
Правила хромосом.
- Видовое постоянство числа хромосом;
- Парности хромосом;
- Индивидуальность хромосом;
- Непрерывность хромосом – способны к самоудвоению (хромосома происходит от хромосомы);
В кариотипе выделяют аутосомы (одинаковые для обоих полов) и половые хромосомы. Или 22 пары.
Метод изучения кариотипа – кариологический анализ – лежит в основе цитогенетического метода.
Суть – изучение препаратов метафазных хромосом.
№5
Ген – это фрагмент молекулы ДНК, содержащий регуляторные элементы и структурную область, и соответствующий одной единице транскрипции, которая определяет возможность синтеза полипептидной цепи или молекулы РНК.
Ген прокариот называется опероном, в его состав входят два основных участка:
- регуляторный (неинформативный),
- структурный (информативный).
У прокариот на долю регуляторных элементов приходится около 10 %, структурных – 90 %.
Структурная область генов прокариот (единица транскрипции) может быть представлена одним кодирующим участком, который называется цистроном, либо несколькими кодирующими участками (полицистронная единица транскрипции).
В структурной зоне закодирована информация о последовательности аминокислот в виде генетического кода. Со структурной области считывается мРНК. При наличии у прокариот полицистронной единицы транскрипции на одном структурном участке одновременно может синтезироваться несколько разновидностей мРНК.
К регуляторным элементам генов прокариот относятся участки, управляющие работой гена:
- промотор,
- оператор,
- терминатор.
Промотор определяет начало транскрипции (участок инициации). С промотором соединяется фермент РНК-полимераза, осуществляющий синтез мРНК.
СТРОЕНИЕ ГЕНА ПРО- И ЭУКАРИОТ
Другой элемент, управляющий процессом транскрипции, – оператор, который располагается поблизости от промотора или внутри него. Этот участок может быть свободным, тогда РНК-полимераза соединяется с промотором и начинается транскрипция. Если оператор связан с белком-репрессором, РНК-полимераза не может нормально соединиться с промотором, и транскрипция невозможна. Следующий регуляторный элемент – терминатор – находится за структурной областью и содержит сигнальный участок остановки транскрипции.

Механизм функционирования системы регуляции синтеза белка был открыт в 1962 году Жакобом и Моно при исследовании культивирования кишечной палочки в лактозной среде и назван lac-опероном.
Упрощенно этот механизм может быть описан следующим образом. На основе информации гена-регулятора синтезируется белок-репрессор; если он активный, он связывается с геном-оператором, перекрывая путь для РНК-полимеразы – процесс трансляции и последующего синтеза белка выключается (запрещается). Если появляется индуктор (например, лактоза в lac-опероне), он присоединяется с белку-репрессору, приводя его в неактивное состояние. Оператор становится активным и включает процесс считывания информации со структурных генов – разрешает трансляцию. Происходит считывание информации с ДНК, начинается синтез необходимого белка – фермента (например, β-галактозидазы в lac-опероне).
Это только один из возможных механизмов, который называется запрещающей индукцией. Существуют и другие механизмы регуляции синтеза белка: разрешающая индукция, разрешающая и запрещающая репрессия, в которых принимают участие апоиндукторы и корепрессоры.
Строение генов у эукариот намного сложнее. Генетическая система эукариот называется транскриптоном. Транскриптон также состоит из двух частей:
- регуляторной (неинформативной),
- структурной (информативной),
относительная пропорция которых противоположна генам прокариот: на долю регуляторного участка приходится 90 %, структурного – 10 %.
Регуляторный участок представляет собой ряд последовательно расположенных промоторов и операторов и несколько терминаторов. Структурный участок состоит из одной единицы транскрипции и имеет “прерывистое” строение: кодирующие участки (экзоны) чередуются с некодирующими (интронами). Одномоментно на структурном участке у эукариот может синтезироваться только одна молекула мРНК, однако благодаря наличию альтернативного сплайсинга в разнос время (в зависимости от потребности клетки) на одной и той же структурной части могут синтезироваться разные виды мРНК (от одной до нескольких десятков).
Социальные кнопки для Joomla
1. Геном прокариот
Основной чертой молекулярной организации прокариот является отсутствие в их клетках ядра, отгороженного ядерной мембраной от цитоплазмы. Отсутствие ядра является лишь внешним проявлением особой организации генома у прокариот.
Геном прокариот построен очень компактно. Количество некодирующих последовательностей нуклеотидов минимально. Многие механизмы регуляции экспрессии генов, использующиеся у эукариот, никогда не встречаются у прокариот. Простота строения генома прокариот объясняется их упрощенным жизненным циклом.
Ген - единица наследственной информации, занимающая определенное положение в геноме или хромосоме и контролирующая выполнение определенной функции в организме. По результатам исследования прокариот, главным образом Е. сoll, ген состоит из двух основных элементов: регуляторной части и собственно кодирующей части. Регуляторная часть гена обеспечивает первые этапы реализации генетической информации, заключенной в структурной части гена; структурная часть гена содержит информацию о структуре кодируемого данным геном полипептида. Количество некодирующих последовательностей в структурной части гена у прокариот минимально. 5"-конец прокариотического гена имеет характерную организацию регуляторных элементов, особенно на расстоянии 50 - 70 н.п. от точки инициации транскрипции. Этот участок гена называют промотором. Он важен для транскрипции гена, но сам в РНК не транскрибируется. Противоположный 3"-конец - терминаторная область, необходимая для тер-минации транскрипции. В РНК он также не транскрибируется. Транскрипция начинается со стартовой точки (+1).
Последовательности ДНК, являющиеся сигналами остановки транскрипции, находятся на 3"-конце гена и называются транскрипционнымитерминаторами. Они содержат последовательности, которые в транскрибируемой РНК формируют структуру шпильки.
Кроме хромосомы у большинства бактерий существуют другие способные к автономной репликации структуры - плазмиды. Это двуцепочечные кольцевые ДНК размером от 0,1 до 5% размера хромосомы, несущие гены, необязательные для клетки-хозяина, или гены, необходимые лишь в определенной среде. Именно такие внехромосомные элементы и содержат гены, которые придают клеткам наследуемую устойчивость к одному или нескольким антибиотикам. Они получили название факторов резистентности, или К-факторов. Другие плазмиды определяют болезнетворность патогенных бактерий, например патогенных штаммов Е. соli, возбудителей чумыи столбняка. Третьи - определяют способность почвенных бактерий использовать необычные источники углерода, например углеводороды нефти.
^ 2. Геном эукариот
Для клеток эукариот характерно наличие оформленного ядра. Информационной макромолекулой их генома является ДНК, которая неравномерно распределена по нескольким хромосомам в виде комплексов с многочисленными белками. Однако генетическую информацию в клетках содержат не только хромосомы ядра. Жизненно важная генетическая информация заключена и во внехромосомных молекулах ДНК. У эукариот - это ДНК хлоропластов, митохондрий и других пластид. Под геномом эукариотического организма в настоящее время понимают суммарную ДНК гаплоидного набора хромосом и каждого из внехромосомных генетических элементов, содержащуюся в отдельной клетке зародышевой линии многоклеточного организма.
Геном эукариот существенно отличается от генома прокариот по ряду признаков, среди которых необходимо отметить его избыточность. Эукариотическая клетка содержит во много раз больше генов, чем прокариотическая. Повышенное содержание ДНК в геноме эукариот нельзя объяснить лишь увеличением потребности этих организмов в дополнительной генетической информации в связи с усложнением организации, поскольку большая часть их геномной ДНК, как правило, представлена некодирующими последовательностями нуклеотидов. Феномен значительной избыточности генома эукариот в отношении некодирующих последовательностей нуклеотидов известен под названием «парадокса С».
Эукариотический ген можно рассматривать как совокупность сегментов ДНК, которые вместе составляют экспрессируемую единицу, ответственную за образование специфического функционального продукта - либо молекулы РНК, либо полипептида.
К сегментам ДНК, составляющим ген, относятся следующие элементы:
1.
Единица транскрипции – это участок ДНК, кодирующий
первичный транскрипт. Он включает: а) последовательность, которая обнаруживается в зрелых функциональных молекулах РНК; б) интроны (для мРНК); в) промежуточные последовательности - спейсеры (для рРНК). Интроны и спейсеры удаляются в
ходе процессинга первичных транскриптов; г) 5"- и 3"-нетранслируемые последовательности (5"-НТП и З"-НТП).
2.
Минимальные последовательности, необходимые для начала
транскрипции (промотор) и конца транскрипции (терминатор).
3.
Последовательности, регулирующие частоту инициации транскрипции, ответственные за индуцибельность и репрессию транскрипции, а также клеточную, тканевую и временную специфичность транскрипции. Они разнообразны по строению, положению и функциям. К их числу относятсяэнхансеры
и сайленсеры
- это последовательности ДНК, расположенные в
тысячах пар нуклеотидов от промотора эукариотического гена и
оказывающие дистанционное влияние на его транскрипцию.
В отличие от прокариотических генов, почти всегда коллинеарных своим РНК, многие гены эукариот имеют мозаичное строение. Под мозаичностью в данном случае подразумевается чередование кодирующих (экзоны) и некодирующих (вставочные последовательности, или интроны) последовательностей в пределах единицы транскрипции. Интроны чаще всего встречаются в генах, кодирующих белки.
Существенную часть генома эукариот (10 - 30%) составляют повторяющиеся последовательности, имеющие определенную структурную организацию и способные перемещаться в геноме как в пределах одной хромосомы, так и между хромосомами. Они получили название подвижных генетических элементов.
Различают два основных класса подвижных генетических элементов: транспозоны и ретротранспозоны. Такая классификация основана на молекулярных механизмах, с помощью которых перемещаются эти элементы.
^ 3. Геномы органелл эукариот: ДНК митохондрий и хлоропластов
Существуют два типа цитоплазматических ДНК: одни находятся в митохондриях эукариот, другие -в хлоропластах зеленых растений и водорослей. Как и все цитоплазматические элементы, они наследуются по материнской линии, а не по законам Менделя! Большая часть белков этих органелл, закодированная в ядерной ДНК, синтезируется в цитоплазме и затем переходит в органеллу. Однако некоторые белки митохондрий и хлоропластов и все их РНК кодируется в ДНК самих органелл и в них же синтезируются. Таким образом, органеллы - это результат объединенных усилий двух геномов и двух трансляционных аппаратов. РНК-компоненты рибосом органелл, а также тРНК, использующиеся при трансляции, кодируются геномами митохондрий и хлоропластов.
Размеры генома хлоропластов у всех исследованных организмов сходны, тогда как митохондриальные геномы у растений намного больше, чем у животных.
Все митохондрии и хлоропласты содержат по несколько копий собственной геномной ДНК. Эти молекулы ДНК обычно распределены в виде отдельных групп в матриксе митохондрий и в строме хлоропластов, где они прикреплены к внутренней мембране. Способ упаковки ДНК неизвестен. По структуре геном более сходен с бактериальным геномом: например, как и у бактерий, у них нет гистонов.
Гены прокариот состоят из двух основных элементов: регуляторной части и собственно кодирующей части(рис. 27). Регуляторная часть обеспечивает первые этапы реализации генетической информации, а кодирующая часть содержит информацию о структуре полипептида, тРНК, рРНК. У прокариот структурные гены, кодирующие белки одноо метаболического пути, часто бывают объединены и называются опероном . Так, например, в лактозном опероне E. coli содержится 3 структурных гена. Для биосинтеза аминокислоты гистидин требуется 9 ферментов и ее оперон содержит 9 структурных генов.
Гены, кодирующие белки, обычно содержат на 5"- и 3"- концах гена или оперона нетранслируемые последовательности (5" – НТП и 3" – НТП ), которые играют важную роль в стабилизации иРНК. Гены тРНК и рРНК отделены друг от друга спейсерами (от англ. – spacer – распорка), т.е. последовательностями, которые вырезаются в ходе их созревания (процессинга)(рис. 27).
(А. С. Коничев, Г. А. Севастьянова, 2005, с. 157)
Гены эукариот имеют более сложное строение. В 1978г. У. Гильберт предположил: эукариотический геном состоит из модульных единиц, что позволяет «смешивать» и «сочетать» части. Онна основании анализа многих работ предложил модель мозаичного (интронно-экзонного) строения гена эукариот (28).Интроны – это некодирующие последовательности, они не входят в состав зрелых РНК.
Экзоны – это последовательности участвующие в образовании зрелых РНК. Они могут быть как кодирующие так и некодирующие. Наследственная информация экзонов реализуется в синтезе определенных белков, а роль интронов до конца еще не выяснена.
Возможное значение интронов:
1. Интроны снижают частоту мутаций, соотношение интронов и экзонов у человека 3:2.
2. Интроны поддерживают структуру ДНК, т.е. играют конститутивную роль.
3. Интроны необходимы для процесса созревания иРНК. Без интронов нарушен выход иРНК в цитоплазму. При введение в ядро искусственной иРНК без интронов, она остается в ядре и в цитоплазму не выходит.
4. В последние годы четко установлено, что некоторые интроны кодируют белки – ферменты, которые их вырезают.
5. Превращаются в малые ядерные РНК (мяРНК).
(А. С. Коничев, Г. А. Севастьянова, 2005 г., с. 157)
Гены высших организмов чаще оказываются прерывистыми, но есть и непрерывистые, например, гены интерферонов, гены гистонов. Степень прерывистости может быть различной – от одного интрона как у гена актина до нескольких десятков, как у гена коллагена(рис.29).
Рис. 29. Карты некоторых прерывистых генов. Жирные линии – экзоны, тонкие - интроны (А. С. Коничев, Г. А. Севастьянова, 2005 г., с. 158)
Длина интронов часто оказывается длинней экзонов: 5 – 20 тыс. и 1 тыс. соответственно. Прерывистость гена считалось достоянием эукариот. Но в 1983г. группа ВЕЗЕ (США) обнаружила их у некоторых археобактерий. Интроны содержаться во всех типах РНК, Интроны в составе иРНК вырезаются при участии мяРНП, которые образуют с интроном сплайсосому. При помощи сплайсосом, узнается начало и конец интрона, их концы соединяются в цепи РНК и интрон вырезается (рис.32).
Эволюционное возникновение мозаичной (итрон – экзонной) структуры генов эукариот в настоящее время не находит объяснения. С точки зрения У. Гильберта появление интронов обеспечило возможность обмена экзонами между неродственными генами. В результате это привело к появлению белков с новыми функциями (гипотеза позднего возникновения интронов). По другой гипотезе интроны это эволюционные реликты, они были частью гигантских генов. Прокариоты являются эволюционным тупиком т.к. не содержат интронов.
Вам также будет интересно:

Взаимодействие
тел
«Ключом
ко всякой науке является вопросительный знак»
Оноре
де...

Имена советского происхождения - личные имена, бытующие в языках народов бывшего СССР,...

Профессиональный стресс
- это напряженное состояние работника, возникающее у него при...

ервоначальный успех в войне был на стороне Англии, одержавшей крупные победы над...

Раздел 1 МЕХАНИКА
Глава 1: О с н о в ы к и н е м а т и к и
Механическое движение....

