





Почему один ген является доминантным, а другой рецессивным? Мутантная ДНК, не кодирующая никакой белок или кодирующая белок, лишенный активности, не будет эксп-рессироваться, т.е. представляет собой рецессивный ген. У гетерозиготных индивидуумов нормальный ген направляет синтез нормального белка, а рецессивный ген никакого вклада в синтез функционального белка не вносит. Такой индивидуум имеет нормальный фенотип; нормальный ген в этом случае доминирует. У индивидуума, гомозиготного по данному рецессивному гену, соответствующий белок не образуется, а поэтому нормальная экспрессия данного признака невозможна. Рецессивный фенотип - это отсутствие нормального признака. Так, карликовость - результат «отсутствия ростовых веществ».
Если белок, о котором идет речь, необходим для жизни данного организма, то организм, неспособный образовать активную форму этого белка, обречен на гибель, и дефектный ген называется в этом случае летальным геном. Доминантные летальные гены существуют, но в большинстве случаев они быстро элиминируются, потому что вызывают гибель несущего их организма. Рецессивные же летальные гены могут и не причинять вреда гетерозиготному индивидууму, поэтому они могут передаваться следующим поколениям и распространяться в популяции весьма широко. Подсчитано, что в среднем человек гетерозиготен приблизительно по 30 летальным рецессивным генам. Это больше, чем у многих других организмов, и отчасти именно этим объясняется тот факт, что у людей среди потомков от браков между близкими родственниками процент летальных наследственных признаков выше, нежели при близкородственных скрещиваниях у большинства других видов.
Если одна копия «нормального» гена не обеспечивает образования его белкового продукта в количестве, достаточном для нормального функционирования организма, то по данному гену наблюдается неполное доминирование. В таких случаях гетерозигота отличается по фенотипу от обеих гомозигот. У человека, например, имеется летальный ген, обусловливающий в гетерозиготном состоянии брахидактилию, т.е. укорочение средней фаланги пальцев; палец при этом выглядит так, как будто в нем не три, а две фаланги. У гомозигот этот ген является причиной аномального развития скелета. Дети, гомозиготные по этому гену, рождаются без пальцев и с другими нарушениями в развитии скелета, приводящими к смерти в раннем возрасте.
В тех случаях, когда брахидактилией страдают оба родителя, можно ожидать, что из каждых четырех детей один будет гомозиготен по этому летальному гену и умрет в раннем детстве, два будут гетерозиготны, т.е. будут страдать брахи-дактилией, и один будет нормальным.
Широко известный ген человека, который в гомозиготном состоянии часто оказывается летальным, ответствен за серповидноклеточную анемию. Соответствующий нормальный ген кодирует одну из полипептидных цепей (бета-цепь) гемоглобина-белка, функция которого заключается в переносе кислорода; этот.белок находится в эритроцитах и обусловливает их красный цвет. У мутантного гена один из нуклеотидов, присутствующих в нормальной ДНК, заменен другим, в результате чего и в бета-цепи гемоглобина одна из аминокислот заменена другой.
Это изменение, казалось бы столь ничтожное, влечет за собой роковые последствия. При аномально низком содержании кислорода молекулы гемоглобина в эритроцитах, содержащих гемоглобин S, агрегируют, образуя волокна. Вследствие этого эритроциты деформируются, приобретая серповидную форму. Такие эритроциты застревают в мелких кровеносных сосудах, и кровоснабжение участков, обслуживаемых этими сосудами, нарушается. Нарушение кровоснабжения вызывает разнообразные симптомы: чувство усталости, судороги, раздражительность.
Ген серповидноклеточности и соответствующий нормальный ген кодоминантны, так что у гетерозиготных индивидуумов синтезируются и нормальные и мутантные бета-цепи. Эритроциты таких людей приобретают серповидную форму лишь при крайне низком содержании кислорода, например, на больших высотах. Без специальных тестов такие гетерозиготные индивидуумы могут и не знать о том, что они являются носителями гена серповидноклеточности. Гомозиготные индивидуумы страдают гораздо сильнее, потому что у них дефектны все бета-цепи.
Индивидуумов, гетерозиготных по тому или иному генетическому нарушению, называют носителями, а гомозиготных- больными. Про людей, гетерозиготных по гену серповидноклеточности, иногда говорят, что они больны серповидноклеточной анемией. Это неверно, ибо это предполагает, что носитель данного гена менее жизнеспособен, нежели нормальный гомозиготный индивидуум. Между тем в действительности это обычно не так.
Около 90% индивидуумов, гомозиготных по гену серповиднооклеточности, умирает в раннем возрасте. Можно было бы ожидать, что такой летальный ген будет быстро элиминироваться естественным отбором, поскольку гомозиготы по этому гену умирают слишком рано и не могут передать его потомству. Однако в тропической Африке на весьма обширных пространствах от 20 до 40% людей гетерозиготны по этому гену. В пятидесятых годах исследователи обратили внимание на один примечательный факт: оказалось, что эти люди живут в местах где распространена особо вирулентная форма малярии.
Болезнь Тея-Сакса, проявляющаяся в нарушении развития головного мозга и приводящая к смерти в возрасте около четырех лет, обусловлена наличием рецессивного летального гена У индивидуумов, гомозиготных по этому гену, отсутствует фермент; ответственный за метаболические превращения определенного липида в клетках мозга. Без этого фермента липид накапливается, и клетки утрачивают способность нормально функционировать. Лечить эту болезнь пока не умеют. Известно, что среди еврейского населения Восточной Европы из каждых 30 человек один гетерозиготен по данному гену. Однако в США приблизительно одна треть всех случаев болезни Тея-Сакса приходится на долю детей из нееврейских семей.
Среди белого населения США чаще всего встречается летальный ген, обусловливающий кистозный фиброз - болезнь, при которой бронхиолы, атакже протоки поджелудочной железы и печени заполняются густой слизью. Жертвы кистозного фиброза умирают обычно от инфекционных респираторных заболеваний в детском и юношеском возрасте, не доживая до 20 лет. Предполагают, что ген, ответственный за кистозный фиброз, кодирует один из ферментов цепи переноса электронов.
Изменение расщепления по фенотипу в отношении 3:1 во втором поколении моногибридного скрещивания бывает связано с разной жизнеспособностью зигот Ф2. Разная жизнеспособность зигот может быть обусловлена наличием летальных генов . Летальным называется ген, вызывающий нарушения в развитии организма, что приводит его к гибели или уродству.
Изучение врожденных аномалий показало, что при разных летальных генах гибель особей бывает различной и может происходить на разных стадиях развития. По классификации, предложенной Розенбауэром (1969), гены, вызывающие гибель 100 % особей до достижения ими половой зрелости, называются летальными , более 50 % -сублетальными (полулетальными) и менее 50 % - субвитальными .
Однако следует отметить, что разделение это до некоторой степени условно и иногда не имеет четких границ. Примером может служить сцепленная с полом голость у кур. Почти половина голых цыплят гибнет в последние 2-3 дня инкубации. Из числа вылупившихся около половины цыплят гибнет до 6-недельного возраста, если их выращивают при температуре 32-35 °С. Но если температура в брудерах будет повышена на 5,5 °С, то погибнет значительно меньше голых цыплят. В 4-5 месяцев у голых цыплят вырастает редкое оперение, и они уже в состоянии переносить довольно низкие температуры. В естественных условиях эта мутация, по-видимому, будет летальной и приведет к 100%-ной гибели птиц. Приведенный пример показывает, что характер проявления полулетального гена в значительной мере может зависеть от условий среды.
Летальные гены могут быть:
- доминантными,
- рецессивными.
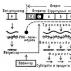
В числе первых летальных факторов был открыт аллель, обусловливающий желтую окраску мышей. Ген желтой окраски является доминантным (Y). Его рецессивный аллель (у) в гомозиготном состоянии обусловливает появление черной окраски. Скрещивание желтых мышей между собой давало в потомстве две части особей желтых и одну часть черных, т. е. получалось расщепление 2:1, а не 3:1, как следовало из правила Менделя. Оказалось, что все взрослые мыши гетерозиготны (Yy). При скрещивании между собой они должны были дать одну часть гомозиготного потомства по желтой окраске (IT), но оно погибает еще в эмбриональном периоде, две части гетерозигот (Yy) будут желтыми и одна часть гомозигот по рецессивному признаку (уу) будет черной. Таким же образом наследуется серая окраска шерсти у каракульских овец (сокольских, малич и др.), платиновая окраска у лисиц, распределение чешуи у линейных карпов и т. д.
Летальные гены в большинстве случаев рецессивны и поэтому долгое время могут находиться в скрытом состоянии. Такие летальные гены оказывают отрицательное воздействие в гомозиготе (рецессивной), в гетерозиготе они безвредны.
Совершенно здоровое и нормальное по фенотипу животное может быть носителем летального гена, действие которого обнаруживается только при переходе в гомозиготное состояние . В гомозиготное состояние летальные гены чаще всего переходят при родственном спаривании. В практике животноводства при разведении лошадей был случай гибели 25 жеребят на 2-4-й день после рождения от уродства прямой кишки - отсутствия заднепроходного отверстия (Atresia ani). Оказалось, что все жеребцы и кобылы, от которых рождались такие ненормальные жеребята, происходили от одного жеребца. Он был гетерозиготен по летальному гену (LI ). Вначале этот жеребец при скрещивании с нормальными кобылами (LL) дал потомство, по фенотипу нормальное, но по генотипу половина потомков была благополучной (LL), a половина - гетерозиготной (LI), несущей рецессивный задаток (0 летального гена. При родственном спаривании гетерозиготных животных (Ы х Ы) появилась часть жеребят, гомозиготных по летальному гену (II), с уродством прямой кишки. Они все погибли.
Изменение расщепления по фенотипу в отношении 3:1 во втором поколении моногибридного скрещивания бывает связано с разной жизнеспособностью зигот Ф2. Разная жизнеспособность зигот может быть обусловлена наличием летальных генов. Летальным называется ген, вызывающий нарушения в развитии организма, что приводит его к гибели или уродству.
Изучение врожденных аномалий показало, что при разных летальных генах гибель особей бывает различной и может происходить на разных стадиях развития.
По классификации, предложенной Розенбауэром (1969), гены, вызывающие гибель 100 % особей до достижения ими пЪловой зрелости, называются летальными, более 50 % --сублетальными (полулетальными) и менее 50 % -- субвитальными. Однако следует отметить, что разделение это до некоторой степени условно и иногда не имеет четких границ. Примером может служить сцепленная с полом голость у кур. Почти половина голых цыплят гибнет в последние 2--3 дня инкубации. Из числа вылупившихся около половины цыплят гибнет до 6-недельного возраста, если их выращивают при температуре 32--35 "С. Но если температура в брудерах будет повышена на 5,5 °С, то погибнет значительно меньше голых цыплят. В 4--5 мес у голых цыплят вырастает редкое оперение и они уже в состоянии переносить довольно низкие температуры.
В естественных условиях эта мутация, по-видимому, будет летальной и приведет к 100%-ной гибели птиц. Приведенный пример показывает, что характер проявления полулетального гена в значительной мере может зависеть от условий среды.
Летальные гены могут быть доминантными и рецессивными. В числе первых летальных факторов был открыт аллель, обусловливающий желтую окраску мышей. Ген желтой окраски является доминантным (Y). Его рецессивный аллель (у) в гомозиготном состоянии обусловливает появление черной окраски. Скрещивание желтых мышей между собой давало в потомстве две части особей желтых и одну часть черных, т. е. получалось расщепление 2:1, а не 3:1, как следовало из правила Менделя. Оказалось, что все взрослые мыши гетерозиготны (Yy). При скрещивании между собой они должны были дать одну часть гомозиготного потомства по желтой окраске (IT), но оно погибает еще в эмбриональном периоде, две части гетерозигот (Yy) будут желтыми и одна часть гомозигот по рецессивному признаку (уу) будет черной. Схема скрещивания выглядит так:
Таким же образом наследуется серая окраска шерсти у каракульских овец (сокольских, малич и др.), платиновая окраска у лисиц, распределение чешуи у линейных карпов и т. д.
Летальные гены в большинстве случаев рецессивны и поэтому долгое время могут находиться в скрытом состоянии.
Совершенно здоровое и нормальное по фенотипу животное может быть носителем летального гена, действие которого обнаруживается только при переходе в гомозиготное состояние. В гомозиготное состояние летальные гены чаще всего переходят при родственном спаривании. В практике животноводства при разведении лошадей был случай гибели 25 жеребят на 2--4-й день после рождения от уродства прямой кишки -- отсутствия заднепроходного отверстия (Atresia ani). Оказалось, что все жеребцы и кобылы, от которых рождались такие ненормальные жеребята, происходили от одного жеребца. Он был гетерозиготен по летальному гену (LI). Вначале этот жеребец при скрещивании с нормальными кобылами (LL) дал потомство, по фенотипу нормальное, но по генотипу половина потомков была благополучной (LL), a половина -- гетерозиготной (LI), несущей рецессивный задаток (0 летального гена. При родственном спаривании гетерозиготных животных (Ы х Ы) появилась часть жеребят, гомозиготных по летальному гену (II), с уродством прямой кишки. Они все погибли.
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РФ
Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования
Пермская государственная сельскохозяйственная академия имени академика Д.Н. Прянишникова
Кафедра: кормление и разведения сельскохозяйственных животных
КОНТРОЛЬНАЯ РАБОТА
по дисциплине: генетика
Выполнила студентка 1 курса
специальности «Ветеринария»
Каменских Ксения Алексеевна
Проверил: старший преподаватель
Пьянкова Светлана Юрьевна
1. Введение
гибридологический генетический аномалия летальный
Генетика представляет собой одну из основных, наиболее увлекательных и вместе с тем сложных дисциплин современного естествознания. Место генетики среди биологических наук и особый интерес к ней определяются тем, что она изучает основные свойства организмов, а именно наследственность и изменчивость.
В результате многочисленных - блестящих по своему замыслу и тончайших по исполнению - экспериментов в области молекулярной генетики современная биология обогатилась двумя фундаментальными открытиями, которые уже нашли широкое отражение в генетике человека, а частично и выполнены на клетках человека.
Это показывает неразрывную связь успехов генетики человека с успехами современной биологии, которая все больше и больше становится связана с генетикой.
Первое - это возможность работать с изолированными генами. Она получена благодаря выделению гена в чистом виде и синтезу его. Значение этого открытия трудно переоценить. Важно подчеркнуть, что для синтеза гена применяют разные методы, т.е. уже имеется выбор, когда речь пойдет о таком сложном механизме как человек.
Второе достижение - это доказательство включения чужеродной информации в геном, а также функционирования его в клетках высших животных и человека. Материалы для этого открытия накапливались из разных экспериментальных подходов.
Прежде всего, это многочисленные исследования в области вирусогенетической теории возникновения злокачественных опухолей, включая обнаружение синтеза ДНК на РНКматрице. Кроме того, стимулированные идеей генетической инженерии опыты с профаговой трансдукцией подтвердили возможность функционирования генов простых организмов в клетках млекопитающих, включая клетки человека. Без преувеличения можно сказать, что, наряду с молекулярной генетикой, генетика человека относится к наиболее прогрессирующим разделам генетики в целом. Ее исследования простираются от биохимического до популяционного, с включением клеточного и организменного уровней.
2. Методы генетически х исследований : гибридологический, генеалогический, популяционный, феногенетический, рекомбинационный, мутационный, цитогенетический, статистический
Для познания закономерностей наследования признаков и их изменчивости генетика использует ряд методов. Основным методом является гибридологический. При этом методе для выявления закономерностей наследования того или иного признака проводится скрещивание особей, различающихся по этому признаку, и изучается полученное потомство в первом и последующих поколениях. Гибридологический метод впервые в своих исследованиях удачно применил Г. Мендель.
Генеалогический метод является одним из вариантов гибридологического. Наследование признака при этом изучают путем анализа передачи его потомству в целых семьях или родственных группах животных, для чего составляют родословные на несколько поколений предков, отдельных особей и целых семей. Генеалогический метод имеет большое значение при изучении наследственности человека и медленно плодящихся животных, к которым обычный гибридологический метод или не применим, или требует продолжительного времени для получения результатов опыта.
Частным случаем гибридологического метода является рекомбинационный метод, основанный на явлении кроссинговера. Его широко используют для составления генетических карт, создания рекомбинантных молекул ДНК, содержащих генетические системы различных организмов.
Цитологический метод применяют при изучении наследственности на уровне клетки и хромосом. Установлено, что многие дефекты и нарушения в организме связаны с изменениями в числе и структуре хромосом. Поэтому при диагностике некоторых наследственных заболеваний человека и животных широко используется цитологический метод.
Биохимический метод используется в генетике для более глубокого анализа нарушений в обмене веществ и их строении. Этот метод используется при манипуляциях на уровне ДНК в генной инженерии.
Популяционно-статистический метод используется для изучения явлений наследственности в популяциях. Этот метод дает возможность установить частоту доминантных и рецессивных аллелей, определяющих тот или иной признак, частоту доминантных и рецессивных гомозигот и гетерозигот, динамику генетической структуры популяций под влиянием мутаций, изоляции и отбора. Широко используется в современной селекции животных.
Феногенетический метод применяется для установления степени влияния генов и факторов внешней среды на развитие признаков организма. При использовании этого метода изучаются особи с разной наследственностью или находящиеся в разных условиях среды.
Мутационный метод (мутагенез) позволяет установить характер влияния мутагенных факторов на генетический аппарат клетки, ДНК, хромосомы, на изменения признаков или свойств. Мутагенез используют в селекции сельскохозяйственных растений, в микробиологии для создания новых штаммов бактерий. Он нашел применение в селекции тутового шелкопряда.
3 . Летальные гены и их действие. Назовите признаки , детерминируемые летальными генами у разных видов сельскохозяйственных животных и покажите их на схемах скрещиваний особенности их наследования
Изменение расщепления по фенотипу в отношении 3:1 во втором поколении моногибридного скрещивания бывает связано с разной жизнеспособностью зигот Ф2. Разная жизнеспособность зигот может быть обусловлена наличием летальных генов. Летальным называется ген, вызывающий нарушения в развитии организма, что приводит его к гибели или уродству.
Изучение врожденных аномалий показало, что при разных летальных генах гибель особей бывает различной и может происходить на разных стадиях развития.
По классификации, предложенной Розенбауэром (1969), гены, вызывающие гибель 100 % особей до достижения ими пЪловой зрелости, называются летальными, более 50 % --сублетальными (полулетальными) и менее 50 % -- субвитальными. Однако следует отметить, что разделение это до некоторой степени условно и иногда не имеет четких границ. Примером может служить сцепленная с полом голость у кур. Почти половина голых цыплят гибнет в последние 2--3 дня инкубации. Из числа вылупившихся около половины цыплят гибнет до 6-недельного возраста, если их выращивают при температуре 32--35 "С. Но если температура в брудерах будет повышена на 5,5 °С, то погибнет значительно меньше голых цыплят. В 4--5 мес у голых цыплят вырастает редкое оперение и они уже в состоянии переносить довольно низкие температуры.
В естественных условиях эта мутация, по-видимому, будет летальной и приведет к 100%-ной гибели птиц. Приведенный пример показывает, что характер проявления полулетального гена в значительной мере может зависеть от условий среды.
Летальные гены могут быть доминантными и рецессивными. В числе первых летальных факторов был открыт аллель, обусловливающий желтую окраску мышей. Ген желтой окраски является доминантным (Y). Его рецессивный аллель (у) в гомозиготном состоянии обусловливает появление черной окраски. Скрещивание желтых мышей между собой давало в потомстве две части особей желтых и одну часть черных, т. е. получалось расщепление 2:1, а не 3:1, как следовало из правила Менделя. Оказалось, что все взрослые мыши гетерозиготны (Yy). При скрещивании между собой они должны были дать одну часть гомозиготного потомства по желтой окраске (IT), но оно погибает еще в эмбриональном периоде, две части гетерозигот (Yy) будут желтыми и одна часть гомозигот по рецессивному признаку (уу) будет черной. Схема скрещивания выглядит так:
Таким же образом наследуется серая окраска шерсти у каракульских овец (сокольских, малич и др.), платиновая окраска у лисиц, распределение чешуи у линейных карпов и т. д.
Летальные гены в большинстве случаев рецессивны и поэтому долгое время могут находиться в скрытом состоянии.
Совершенно здоровое и нормальное по фенотипу животное может быть носителем летального гена, действие которого обнаруживается только при переходе в гомозиготное состояние. В гомозиготное состояние летальные гены чаще всего переходят при родственном спаривании. В практике животноводства при разведении лошадей был случай гибели 25 жеребят на 2--4-й день после рождения от уродства прямой кишки -- отсутствия заднепроходного отверстия (Atresia ani). Оказалось, что все жеребцы и кобылы, от которых рождались такие ненормальные жеребята, происходили от одного жеребца. Он был гетерозиготен по летальному гену (LI). Вначале этот жеребец при скрещивании с нормальными кобылами (LL) дал потомство, по фенотипу нормальное, но по генотипу половина потомков была благополучной (LL), a половина -- гетерозиготной (LI), несущей рецессивный задаток (0 летального гена. При родственном спаривании гетерозиготных животных (Ы х Ы) появилась часть жеребят, гомозиготных по летальному гену (II), с уродством прямой кишки. Они все погибли.
4. П енетрантность и экспрессивность при наследовании аномалий
При доминантном типе наследования наблюдаются случаи, когда от аномального родителя рождается нормальный потомок, который, в свою очередь, дает аномальный приплод. Из этого ясно, что «нормальное» животное было гетерозиготным носителем аномального гена, который, однако, не проявился в его фенотипе. Доминантные признаки, которые не всегда проявляются в фенотипе, получили название признаков с неполной пенетрантностью.
Различают признаки с полной и неполной, средней или низкой степенью пенетрантности, выраженные в процентах или долях единицы. К доминантным аномалиям с неполной пенетрантностью гена можно отнести анкилоз всех суставов в сочетании с «волчьей пастью», летальную аномалию «баранья голова» у крупного рогатого скота и др. У коз комолость обусловлена рецессивным геном рр и сочетается с нарушением плодовитости вследствие закупорки придатка семенника. При скрещивании комолых родителей рождается около 2€Г % козлов, полностью непригодных для воспроизводства.
Пенетрантность стерильности колеблется от 0,42 до 0,62.
Как уже известно, существует понятие экспрессивности генов. В качестве примеров разной экспрессивности генов можно привести следующие. Так, адактилия у крупного рогатого скота варьирует от частичного до полного отсутствия фаланг конечностей. При исследовании пупочных грыж у телят мы обнаружили значительные различия в диаметре грыжевого отверстия. При изучении нарушения координации движений у телят черно-пестрой и симментальской пород мы наблюдали заметную разницу во внешнем проявлении аномалии -- от дрожания головы и покачивающейся ходьбы до резко выраженной несогласованности движений головы и конечностей при попытке животных встать. Патологоанатомическими и гистологическими исследованиями установлена различная степень гипоплазии мозжечка у телят, что сочеталось со степенью фенотипического проявления аномалии.
У сельскохозяйственных животных известны десятки аномалий, возникновение которых связано с рецессивными или доминантными мутациями генов. Эти аномалии встречаются в отдельных популяциях с разной частотой, что зависит от скорости мутационного процесса, системы разведения животных и др. Знание конкретных форм врожденных аномалий у животных каждого вида, а также частоты их проявления в отдельных породах необходимо ветеринарным специалистам для селекционной профилактики распространения генетической патологии.
Аномалии у крупного рогатого скота. Биологические особенности данного вида животных -- малоплодие и относительная позднеспелость. Корова обычно приносит одного теленка, который достигает половой и физиологической зрелости только к 1,5 года, так что период между первым отелом матери и дочери в среднем составляет 5 лет. Вследствие этого появление в стаде аномального приплода может существенно снижать уровень воспроизводства и интенсивность племенного отбора поголовья. У крупного рогатого скота изучен широкий спектр врожденных аномалий, детерминированных летальными, полулетальными и субвитальными генами. 46 аномалий включено в Международный список летальных дефектов под шифром А (табл.).
Список генетически обусловленных аномалий у крупного рогатого скота
|
Признак аномалии |
1 Тип наследования |
|
|
1. Укорочение нижней челюсти |
||
|
2. Укорочение верхней и нижней челюстей (мопсовидность) |
||
|
3. Отсутствие нижней челюсти |
||
|
4. Укорочение верхней челюсти |
||
|
5. «Волчья пасть», «заячья губа» н др. черепно-лицевые дефекты |
||
|
6. Отсутствие носовых отверстий |
||
|
7. «Баранья голова» -- выгнутый, как у барана, лицевой профиль головы |
||
|
8. Мозговая грыжа |
||
|
9. Водянка головного мозга |
||
|
10. Уменьшение размеров глазного яблока |
||
|
11. Отсутствие глазных яблок |
||
|
12. Помутнение хрусталика |
||
|
13. Слепота |
||
|
14. Альбинизм |
||
|
15. Искривление и деформация передних конечностей, неподвижность в запястных суставах |
||
|
16. Контрактура задних конечностей, закрюченность их в сторону брюха - |
||
|
17. Неподвижность конечности (мышечная контрактура) |
||
|
18. Контрактура мышц конечностей и направленность их назад |
||
|
19. Неподвижность (анкилоз) всех суставов, «волчья пасть» |
||
|
20. Искривление и анкилоз передних конечностей |
||
|
21. Отсутствие задних конечностей |
||
|
22. Отсутствие передних н задних конечностей |
||
|
23. Редукция числа позвонков |
||
|
24. Расцепление позвоночника |
||
|
25. Бульдогообразная карликовость |
||
|
26. Пропорциональная карликовость |
||
|
27. Коротконогость в сочетании с дефектами челюсти |
||
|
28. Водянка плода |
||
|
29. Мумификация плода |
||
|
30. Бесшерстность |
||
|
31. Отсутствие кожи на отдельных участках |
||
|
32. Удлинение сроков стельности коров |
||
|
33. Слияние копытец |
||
|
34. Эпилепсия |
||
|
35. Паралич задних конечностей |
||
|
36. Хромота задних конечностей (спастический парез) |
||
|
37. Нарушение координации движений |
||
|
38. Порфирия (повышенная светочувствительность) |
||
|
39- Гиперемия кожи и слизистых оболочек, одышка, наруше ние роста |
||
|
40. Нарушение обмена цинка (дерматиты) |
||
|
41. Отсутствие фермента маннозидазы (агрессивность, летальный исхоп} |
||
|
Тс "* гш HvAU^I 42. Бесхвостость, отсутствие костей таза, аномалии прямой |
||
|
кишки и урогенительной системы |
||
|
43. Пупочная грыжа |
||
|
44. Хрупкость кожи и периферических кровеносных сосудов, |
||
|
отеки кожи |
||
|
45. Скручивание и деформация скелета |
||
|
46. Дефекты морфологического строения спермиев (дефекты |
||
|
акросомы, деформации хвоста и средней части, образова- |
||
|
ние петель, эксцентричное прикрепление хвоста, дезинтег- |
||
|
рация спермиев) |
Обозначения: ар -- аутосомный рецессивный; д -- доминантный; дни -- доминантный с неполной пенетрантностью; спр -- сцепленный с полом рецессивный.
Относительная частота отдельных типов аномалий в каждой породе или популяции может быть различной. В костромской породе, по нашим данным, наиболее часто регистрируется генетическая аномалия головы -- укорочение челюсти (табл. 44), в ярославской породе -- синдактилия, в холмогорской -- контрактуры мышц, в черно-пестрой -- пупочные грыжи. У крупного рогатого скота в Германии наиболее часто (21 %) встречались аномалии центральной нервной системы.
Второе место по частоте регистрации (14 %) занимала комплексная аномалия -- сочетания пупочных грыж с расщеплением брюха и плода в целом.
Частота отдельных форм врожденных аномалий у телят костромской породы (по данным племенного хозяйства за 1969--1982 гг.)
|
Форма аномалии |
аномальных телят |
||
|
1 % от общего поголовья |
|||
|
Укорочение нижней челюсти |
|||
|
Мопсовидность |
|||
|
Водянка плода |
|||
|
«Щучья голова» |
|||
|
Атрезия ануса |
|||
|
Уродства конечностей |
|||
|
Вывернутые лопатки |
|||
|
Искривление костяка |
|||
|
Искривление хвоста |
|||
|
Искривление конечностей |
|||
|
Слабость конечностей |
|||
|
Неклассифицированные уродства |
|||
Частота аномалий, или процентное отношение аномального приплода к общему его числу, в пределах конкретных популяций также может быть самым различным и по средним оценкам не превышать 1 %. Однако этот показатель зависит от полноты и точности регистрации аномалий. Так, в Германии после организации четкого учета сделали вывод, что частота аномалий возросла в несколько раз. Вопрос в другом: все ли аномалии поддаются визуальному наблюдению? Очевидно, не все. Так, в костромской породе средняя частота всех форм аномалий за 12-летний период составила 1,15 %. Частота же общей смертности приплода (абортированные, мертворожденные, аномальные, павшие без видимых дефектов телята) в этом хозяйстве составила 10,2 %. Определенная доля этой смертности также связана с генными мутациями, вызывающими не морфологические дефекты, а нарушение обмена и другие аномалии, выявление которых возможно только специальными методами.
Особую роль в распространении генетических аномалий как у крупного рогатого скота, так и у животных других видов могут сыграть производители. От каждого производителя при искусственном осеменении в год можно получить сотни и тысячи потомков. Так, от одного быка за рубежом получили 100 тыс. телят. Если такой производитель окажется носителем генной мутации, то она быстро распространится в породе. Вот некоторые примеры из многочисленных фактов, описанных в литературе. В результате интенсивного использования быка Принца Адольфа, завезенного в Швецию, и последующего стихийного инбридинга на него частота бесшерстности в отдельных шведских стадах была свыше 5 %. Такая же ситуация сложилась в Швеции после импорта быка Галлуса, который оказался гетерозиготным носителем гена, обусловливающего отсутствие конечностей.
В потомстве отдельных быков черно-пестрой породы и шароле в США и Германии регистрировали случаи рождения карликовых телят с частотой соответственно 23,3 и 22,2 %. В бывш. Чехословакии при обследовании потомков 166 быков-производителей уста-новлеш!, что 43 из них были носителями летальных генов. У одного быка -- носителя доминантной аномалии «заячья губа» -- дефект проявился среди 44 % бычков и 71 % телочек из его потомства.
В костромской породе нами проанализировано распространение укорочения нижней челюсти и мопсовидности через быка
Рис. 1. Схема, иллюстрирующая рецессивное наследование укорочения челюсти в одной линии костромской породы
Бурхана, который сам имел в потомстве дефектных телят; его сыновья, внуки, правнуки, женские потомки также давали аномальный приплод (рис. 57). Большинство телят получено от родственного спаривания и спаривания родителей с нормальным фенотипом при наличии общего предка. Следовательно, можно сделать вывод о рецессивном типе наследования данной аномалии.
Из рисунка видно, что наибольшее количество дефектных телят зарегистрировано в потомстве быка Жетона 3501 (внука быка Бурхана) при использовании его в товарном хозяйстве, где часть коров имела в генотипе такой же рецессивный ген.
Аномалии у свиней. В Международный список летальных дефектов свиней включено 18 генетических аномалий. Основная часть их обусловлена аутосомными рецессивными генами (табл.).
Слисок генетически обусловленных аномалий у свиней
|
Фенотип аномалии 1 Тип няр.ле.лппяния |
||
|
1. Мозговая грыжа |
||
|
2. Паралич задних конечностей |
||
|
3. Отсутствие анального отверстия |
||
|
4. Расщепление нёба («волчья пасть») |
||
|
5. Толстоногость |
||
|
6. Искривление и ригидность конечностей |
||
|
7. Недоразвитие ушных раковин |
||
|
8. Водянка головного мозга |
||
|
9. Отсутствие конечностей |
||
|
10. Дивертикулез подвздошной кишки |
||
|
11. Порфирия |
||
|
12. Отеки на затылке. Зобообразная припухлость шеи |
||
|
13. Гиперкератоз |
||
|
14. Желтуха новорожденных |
||
|
15. Гемофилия |
||
|
16. Укорочение позвоночника, череггао-лицевые дисплазии |
||
|
17. Дефекты кожи |
||
|
18. Агенезия мышц сфинктера заднего прохода |
||
|
19. Укорочение верхней челюсти |
||
|
20. Укорочение нижней челюсти |
||
|
21. Отсутствие нижней челюсти |
||
|
22. Уменьшение размеров глазного яблока |
||
|
23. Водянка глазного яблока |
||
|
24. Циклопия |
||
|
25. Альбинизм |
||
|
26. Эпилепсия и судороги |
||
|
27. Короткохвостость в сочетании с гидроцефалией |
||
|
28. Трехногие поросята |
||
|
29. Полное или частичное отсутствие фаланг |
||
|
30. Искривление задних конечностей кпереди и в сторону |
||
|
31. Пупочная грыжа |
||
|
32. Дефекты акросомы спермиев |
||
|
33. Крипторхизм |
Обозначения: ар-- аугосомный рецессивный; д -- доминантный; нд -- неполный доминантный; п -- полигенный.
Генетические аномалии могут занимать значительное место в патологии свиней. Рассмотрим несколько таких примеров. В Испании при исследовании 23 449 поросят из* 2399 пометов, полученных от хряков пород дюрок, йоркшир, гемпшир и белый честер, выявили соответственно 6,21; 6,02; 9,66; 2?62 % аномальных пометов.
Согласно данным Оливье (1979), у свиней описано 7 генетических аномалий кожного покрова, 17 -- скелета, 3 ~ глаз, 13 -- нервно-мышечных, 6 -- крови, 6 -- гормонально-обменных, 5 -- пищеварительной системы, 9 -- мочеполовой. Основными аномалиями были крипторхизм, грыжи, псевдогермафродитизм и др. Автор исследования считает, что эти аномалии -- результат действия одного гена на разных этапах формирования эмбриона.
В Дании для выяснения характера и частоты возникновения врожденных аномалий в течение двух лет исследовали 6669 павших поросят из 2936 пометов. Различные аномалии выявлены у 1,4 % поросят к числу родившихся, или 6,2 % к числу павших до отъема особей. При патологоанатомическом исследовании у 25,9 % аномальных поросят были обнаружены недоразвитие клапанов, незакрытие анального отверстия, субаортальные стенозы, эктопия сердца и другие дефекты сердечно-сосудистой системы. У 23,4 % поросят найдены различные нарушения развития двигательной системы. Аномалии центральной нервной системы выявлены у 5,9 % поросят, среди них раздвоенный мозг и водянка мозга. Заращение прямой кишки, тонких кишок или их неполное развитие обнаружены у 30 % поросят, а различные грыжи и асцит -- у 6,8 %. «Заячья губа», расщепление нёба, риноцефалит и другие аномалии лицевой части головы найдены у 6,1 %; гермафродитизм, незаращение мочеточника, водянка почек и уретры--у 1,7 % поросят. Указанные аномалии проявлялись в потомстве отдельных производителей при инбридинге, что указывает на наследственный характер их возникновения.
Весьма убедительные доказательства наследственной природы крипторхизма у свиней получили Фридин и Ньюман. По их данным, в Канаде одно-, двухсторонний крипторхизм наблюдается ежегодно у 1--2 % всех хрячков, поступающих на рынок. Авторы скрещивали крипторхов с их матерями и полными сестрами. Потомков от таких скрещиваний спаривали между собой. В результате такого отбора и подбора частота крипторхизма у экспериментальных животных йоркширской породы возросла в среднем^ до 42,9 % и особенно при использовании двух производителей. При обследовании поросят в США за один год было обнаружено около 400 тыс. животных с мошоночной грыжей.
Наблюдения показывают, что причиной нарушения плодовитости у хряков нередко бывает гипоплазия семенников. Частота этой аномалии, по данным исследователей из Германии, составила 19,6 %, 30 таких хряков были оставлены для воспроизводства, каждым из них было покрыто от 4 до 40 маток (всего 439 голов), но лишь четыре из них дали потомство. Анализ показал, что патологические формы спермиев у этих хряков составляют 80--100 %. Все 30 аномальных животных имели общих предков, что указывает на наследственный характер гипоплазии семенников и дефектов спермиогенеза.
Наличие кратерных сосков у свиней -- один из серьезных дефектов, поскольку поросята не получают от них молока. По данным Баварского института животноводства (Германия), частота этой аномалии у немецких ландрасов составила 6,6 %. Как отмечают П. Н. Кудрявцев и др. (МВА), количество свиней с бездействующими кратерными сосками в последние годы увеличилось. Число таких сосков колеблется от 1 до 8. Поросята, которым достаются кратерные соски, погибают.
Кратерность -- признак, обусловленный одним аутосомным рецессивным геном. Экспериментально это проверили П. Н. Кудрявцев с сотр. Предварительно выделив хряков и свинок -- носителей аномалий {кчкч), нормальных, но гетерозиготных особей {Кчкч), дающих в приплоде поросят с кратерностью, и нормальных гомозиготных свиней {КчКч), авторы провели скрещивания между этими группами животных; В первом варианте скрещивали 27 нормальных гомозиготных маток с 15 хряками. Все 258 потомков были нормальными. Во втором варианте, где один из родителей был гомозиготным {КчКч), а другой -- гетерозиготным {Кчкч), все поросята также были нормальными. В третьем» варианте скрещивали 13 гетерозиготных хряков с 16 гетерозиготными матками. Из 168 родившихся поросят 39 (23,2 %) имели кратерные соски. И наконец, в четвертом варианте один из родителей был гомозиготным, а другой -- гетерозиготным. От них было получено 170 поросят, из них 86 (50,5 %) оказались с нормальными сосками и 84 (49,5 %) -- с кратерными. Результаты этого опыта доказывают рецессивный тип наследования кратерности сосков у свиней.
Аномалии у овец. У овец описано около 90 врожденных аномалий. По данным Денниса и Лейпольда, большинство известных генетических дефектов у овец обусловлено моногенным аутосомным рецессивным типом наследования (табл. 46). Наиболее часто у этого вида животных встречаются черепно-лицевые дефекты, особенно агнатия, а также искривление передних конечностей, микроагнатия, гермафродитизм, крипторхизм, гипос-падиаз, прогнатия, атрезия ануса, микротия, энтропия, тортико-лис, полителия, артрогрипоз. Анализ показал, что 55,4 % дефектов относились к мышечно-скелетной системе, 12,7 -- к пищеварительной, 9,7 -- к сердечно-сосудистой, 7,1 -- к урогени-тальной, 6 -- к центральной нервной системе, 3,5 -- к аномалии связок, 3,2 -- к брюшной, 1,5 % -- к эндокринной системе. Хотя частота отдельных дефектов и невысока, однако кумулятивный вклад всех аномалий может нанести ущерб хозяйствам.
Список генетически обусловленных аномалий у овец
|
Фенотип аномалии |
Тип наследования |
|
|
1. Отсутствие нижней челюсти и непроходимость пищевода |
||
|
2. Выпадение шерсти, облысение |
||
|
3. Коротконогость |
||
|
4. Мышечная контрактура -- сильно согнутое положение мус- |
||
|
кулатуры конечностей. Слабое развитие. Ягнята обычно мертворожденные |
||
|
5. Уменьшение размеров глазных яблок или их отсутствие -- слепота |
||
|
6. Отсутствие мышечной координации. Некоторые ягнята слепые и глухие |
||
|
7. Неопущение семенников в мошонку (крипторхизм) |
||
|
8. Карликовость |
||
|
9. Недоразвитие ушной раковины и «волчья пасть» |
||
|
10. Летмьная серая окраска |
||
|
11. Летальная мышечная дистрофия. Ягнята рождаются живыми, но погибают вскоре после рождения |
||
|
12. Паралич тазовых конечностей |
||
|
13. Деформация скелета в сочетании с короткой шерстью и грыжами |
||
|
14. Частичное или полное отсутствие фаланг конечностей |
||
|
IS. Светочувствительность, красно-коричневая окраска костей и зубов |
||
|
16. Непроходимость ануса |
||
|
17. Врожденная водянка. Скопление жидкости в подкожной |
||
|
соединительной ткани грудной и брюшной полостей |
||
|
18. Отсутствие дистальных отделов конечностей |
||
|
19. Ватность. Отсутствие ости |
||
|
20. Узкоглазость |
||
|
21. Дистрофия нервных волокон. Нарушение двигательных функций 22. Избыток билирубина |
||
|
23. Коллагенная дисплазия. Нарушение формирования коллагена |
||
|
24. Искривление шеи |
||
|
25. Недоразвитие и отсутствие ушей. Глухота |
||
|
26. Укороченные челюсти |
||
|
27. Судороги головы и шеи, ягнята не могут двигаться |
||
|
28. Одноглазие |
||
|
29. Дефекты наружной кожи, язвы |
||
|
30. Заворот век внутрь глаза |
||
|
31. Пупочные грыжи |
||
|
32. Бесшерстность |
||
|
33. Недоразвитие мужских половых органов |
||
|
34. Уменьшение размеров черепа и головного мозга |
||
|
35. Нарушение развития костяка |
||
|
36. «Волчья пасть» (расщепление нёба) |
||
|
37. Несформированный вырез мошонки -- |
||
|
38. Отсутствие задней части позвоночника -- |
||
|
39. Постоянные пупочные кровотечения -- |
||
|
40. Затянувшаяся беременность ~ |
||
|
41. Ложный гермафродитизм. Бесплодие -- |
||
|
42. Однокопытность. Слияние копытного рога -- |
||
|
43. Уменьшение размеров одного или двух семенников |
Обозначения: ар -- аугосомный рецессивный; д -- доминантный.
В Новой Зеландии, в стране развитого овцеводства, частота летальных дефектов составила около 1 % павших ягнят. В США летальные дефекты в первые 2 нед жизни наблюдались у 11,4 %
Средняя эмбриональная смертность у овец равна 20 %. Это указывает на то, что в этот период могут проявлять свое действие многие неидентифицируемые летальные гены.
Отдельные генетические аномалии у овец могут быть широко распространены. Так, в Болгарии в стадах мериносовых овец наблюдали высокую смертность ягнят в ранний постнатальный период. Она наступала в результате того, что ягнята не получили молока из-за дефектов вымени их матерей: от гипоплазии с небольшими остатками железистой ткани до полного отсутствия ее. Частота этой наследственной аномалии в разных стадах колебалась от 6 до 40 %.
У овец часто наблюдали крипторхизм, который сочетался с таким вторичным половым признаком, как комолость. Комолые бараны имеют низкую плодовитость. Путем жесткого племенного отбора удалось создать тип комолых баранов с нормальной плодовитостью, однако и среди них встречаются аномальные особи.
Аномалии у ипщ. Птицы, прежде всего куры, наиболее изучены в отношении генетики аномалий. В Международный список летальных дефектов включено 45 аномалий у кур, 6 у индеек и 3 у уток Наиболее часто встречаются аномалии клюва (клюв попугая перекрещивающийся клюв). Частота их, по данным Уильяма и др составляет 1,1 % потерь при инкубации яиц кур пород белый леггорн и род-айланд. Аномалии клюва часто встречаются и у уток.
Макс Джиббон и Шейкелферд описали аномалию при скрещивании белых леггорнов с батеркампами и бентамками и последующем разведении «в себе» - полидактилию. Кроме нее у цыплят наблюдали синдактилию и оперенные ноги. Частота синдрома составляла 16,8 %. При скрещивании аномального петуха F> с нормальными по фенотипу курами наблюдали расщепление -- половина нормальных и половина аномальных особей (1:1). Установлено, что данный комплекс признаков контролируется одним аутосомным геном, обладающим полулетальным эффектом, так как эмбриональная и постэмбриональная выживаемость аномальных цыплят очень низка.
Аномалии у лошадей. Из наследственных аномалий у лошадей 10 включены в Международный список летальных дефектов. Среди них 3 аномалии скелета, 2 -- воспроизводительной системы, 2 -- почек и мышц, по одной аномалии кишечника, нервной системы, органов зрения.
У лошадей тяжеловозных пород чаще встречается атрезия ободочной кишки. Распространение этой аномалии было отмечено в потомстве жеребца Суперба першеронской породы. Описана она также у жеребят чистокровной верховой породы остфризского происхождения. У тяжеловозных пород зарегистрирован несовершенный эпителиогенез. У жеребят ольденбургской породы в Германии обнаружена атаксия, названная ольденбургской. Распространилась она в линии 9. Одна из наиболее часто регистрируемых аномалий у лошадей -- пупочная грыжа. Встречается она у легких и тяжеловозных пород.
У лошадей известны некоторые другие генетические и наследственно-средовые аномалии. Так, в США у лошадей нескольких пород наблюдается появление жеребят со своеобразной белой пятнистостью, названной «оверо». При скрещивании лошадей типа «оверо» рождаются жеребята с розовой кожей, у которых наблюдают гипоплазию кишечного тракта и изоэритролиз, а также колики, приводящие к гибели.
В Англии у лошадей чистокровной верховой породы зарегистрированы и исследованы животные с нарушением координации движений -- «болезнь вихляния». Установлена генетическая предрасположенность к этой аномалии.
Доказана наследуемость дерматозов конечностей у лошадей. Пороками с наследственной предрасположенностью являются нередко встречающееся хроническое деформирующее воспаление скакательного сустава -- шпат, так называемая «костыльная нога» у жеребят, а также хроническое асептическое воспаление венечного блока копыт, наблюдаемое в основном у скаковых и беговых лошадей.
ЗАКОН ГОМОЛОГИЧЕСКИХ РЯДОВ В НАСЛЕДСТВЕННОЙ ИЗМЕНЧИВОСТИ
Выдающийся советский ученый академик Н. И. Вавилов при изучении коллекций диких и культурных растений, собранных во время экспедиции в разных странах мира, обнаружил в них много общих наследственных изменений. На основании этих исследований он сформулировал закон гомологических рядов наследственной изменчивости:
1) генетически близкие виды и роды характеризуются сходны ми рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть существование параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и линнеоны, тем полнее сходство в рядах их изменчивости;
2) целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды, составляющие семейство.
Этот закон, как показали дальнейшие исследования ученых, имеет универсальный характер. Обнаружено сходство мутаций не только у растений, но и у животных. Так, были подмечены появления аналогичных форм аномалий у разных видов животных, что указывает на сходство строения многих ферментов и белков и соответственно на сходство генотипов у них. Эти данные подтверждают закон гомологических рядов. Зная формы аномалий у одного вида животных, следует предполагать, что они имеются или могут возникнуть и у другого вида, близкого с первым по происхождению (табл.).
Наследственные дефекты, встречающиеся на аналогичной форме у разных видов домашних животных (по Хатту, 1969; Мейеру, Велюру, 1974)
|
Аномалии систем органов |
Крупный рогатый скот |
|||||||
|
дефектность эпителия |
||||||||
|
отсутствие шерсти или перьев |
||||||||
|
ихтиоз (чешуйчатость кожи) |
||||||||
|
карликовость |
||||||||
|
бульдогообразная карликовость |
||||||||
|
укорочение позвоночника |
||||||||
|
отсутствие конечностей |
||||||||
|
укорочение челюсти |
||||||||
|
расщепление нёба |
||||||||
|
«заячья губа» |
||||||||
|
Нервной системы: |
||||||||
|
паралич задних конечностей |
||||||||
|
контрактура мышц |
||||||||
|
водянка головы |
||||||||
|
мозговая грыжа |
||||||||
|
Внутренних органов: |
||||||||
|
урогенитальные дефекты |
||||||||
|
крипторхизм |
||||||||
|
Аномалии систем органов |
Крупный рогатый скот |
|||||||
|
Обмена веществ и эндокринной системы: порфирия |
Обозначения: р -- рецессивный тип наследования; д -- доминантный; х -- тип наследования точно не установлен.
Размещено на Allbest.ru
...Подобные документы
Рассмотрение процесса разведения и развития сельскохозяйственных животных, наследования хозяйственно-полезных признаков, проведение сравнительной характеристики линий по признакам продуктивности. Совершенствование племенных и продуктивных качеств стада.
курсовая работа , добавлен 25.03.2018
Краткая характеристика различных генетических и паратипических факторов, которые в той или иной степени влияют на уровень продуктивности сельскохозяйственных животных. Гибридизация как скрещивание животных, принадлежащих к разным видам, ее эффективность.
курсовая работа , добавлен 09.07.2013
Аутосомно-доминантные и рецессивные наследственные аномалии у животных. Семь генетических аномалий кожного покрова у свиней. Дерматозы конечностей у лошадей. Генетическая устойчивость животных к болезням. Мастит, туберкулез, бруцеллез, лептоспироз.
презентация , добавлен 23.01.2016
Дикие предки и родичи домашних животных. Изменение животных под влиянием одомашнивания: величина и формы тела, масть и волосяной покров, плодовитость. Характерные признаки доместикации. Изучение происхождения и эволюции сельскохозяйственных животных.
реферат , добавлен 01.03.2015
Санитарные правила искусственного осеменения сельскохозяйственных животных. Клинические методы диагностики беременности и бесплодия животных. Подготовка акушерских инструментов к процессу родовспоможения. Способы гинекологических исследований самок.
контрольная работа , добавлен 02.10.2010
Оценка сельскохозяйственных животных по родословной. Значение конституции и экстерьера при оценке и выборе животных. Способы оценки по экстерьеру и конституции. Методы оценки производителей по качеству потомства в молочном и молочно-мясном скотоводстве.
реферат , добавлен 18.03.2011
Понятие и клиническая картина, факторы риска развития эшерихиоза как остро протекающей зоонозной болезни молодняка животных многих видов. Его симптоматика и закономерности диагностики, схема лечения и прогноз. Характеристика и действие возбудителя.
презентация , добавлен 25.04.2016
Описание последствий скрытых форм витаминной недостаточности (гиповитаминозов) у животных: нарушения воспроизводства, снижения устойчивости к болезням, замедления роста и продуктивности. Организация витаминного питания сельскохозяйственных животных.
реферат , добавлен 14.12.2011
Обзор литературы по теме "Коэффициент наследуемости и повторяемости". Материалы и методика исследований. Анализ результатов исследования. Общие положения и генетические основы отбора. Сущность повторяемости признаков и коэффициента наследуемости.
реферат , добавлен 17.06.2010
Рассмотрение основ спаривания (случки) сельскохозяйственных животных. Изучение основных типов естественного осеменения. Половые рефлексы отдельных вводов животных и их особенности. Оценка результативности и экономической эффективности спаривания.
Изменение расщепления по фенотипу в отношении 3:1 во втором поколении моногибридного скрещивания бывает связано с разной жизнеспособностью зигот Ф2. Разная жизнеспособность зигот мэжет быть обусловлена наличием летальных генов. Летальным называется ген, вызывающий нарушения в развитии организма, что приводит его к гибели или уродству.
Изучение врожденных аномалий показало, что при разных летальных генах гибель особей бывает различной и может происходить на разных стадиях развития.
По классификации, предложенной Розенбауэром (1969), гены, вызывающие гибель 100 % особей до достижения ими пЪловой зрелости, называются летальными, более 50 % -сублетальными (полулетальными) и менее 50 % - субвитальными. Однако следует отметить, что разделение это до некоторой степени условно и иногда не имеет четких границ. Примером может служить сцепленная с полом голость у кур. Почти половина голых цыплят гибнет в последние 2-3 дня инкубации. Из числа вылупившихся около половины цыплят гибнет до 6-недельного возраста, если их выращивают при температуре 32-35 "С. Но если температура в брудерах будет повышена на 5,5 °С, то погибнет значительно меньше голых цыплят. В 4-5 мес у голых цыплят вырастает редкое оперение и они уже в состоянии переносить довольно низкие температуры. В естественных условиях эта мутация, по-видимому, будет летальной и приведет к 100%-ной гибели птиц. Приведенный пример показывает, что характер проявления полулетального гена в значительной мере может зависеть от условий среды.
Летальные гены могут быть доминантными и рецессивными. В числе первых летальных факторов был открыт аллель, обусловливающий желтую окраску мышей. Ген желтой окраски является доминантным (Y). Его рецессивный аллель (у) в гомозиготном состоянии обусловливает появление черной окраски. Скрещивание желтых мышей между собой давало в потомстве две части особей желтых и одну часть черных, т. е. получалось расщепление 2:1, а не 3:1, как следовало из правила Менделя. Оказалось, что все взрослые мыши гетерозиготны (Yy). При скрещивании между собой они должны были дать одну часть гомозиготного потомства по желтой окраске (IT), но оно погибает еще в эмбриональном периоде, две части гетерозигот (Yy) будут желтыми и одна часть гомозигот по рецессивному признаку (уу) будет черной. Схема скрещивания выглядит так:
Таким же образом наследуется серая окраска шерсти у каракульских овец (сокольских, малич и др.), платиновая окраска у лисиц, распределение чешуи у линейных карпов и т. д.
Летальные гены в большинстве случаев рецессивны и поэтому долгое время могут находиться в скрытом состоянии. Совершенно здоровое и нормальное по фенотипу животное может быть носителем летального гена, действие которого обнаруживается только при переходе в гомозиготное состояние. В гомозиготное состояние летальные гены чаще всего переходят при родственном спаривании. В практике животноводства при разведении лошадей был случай гибели 25 жеребят на 2-4-й день после рождения от уродства прямой кишки - отсутствия заднепроходного отверстия (Atresia ani). Оказалось, что все жеребцы и кобылы, от которых рождались такие ненормальные жеребята, происходили от одного жеребца. Он был гетерозиготен по летальному гену (LI). Вначале этот жеребец при скрещивании с нормальными кобылами (LL) дал потомство, по фенотипу нормальное, но по генотипу половина потомков была благополучной (LL), a половина - гетерозиготной (LI), несущей рецессивный задаток (0 летального гена. При родственном спаривании гетерозиготных животных (Ы х Ы) появилась часть жеребят, гомозиготных по летальному гену (II), с уродством прямой кишки. Они все погибли. (Более подробно об аномалиях при летальных генах будет сказано в соответствующей главе.)
Конец работы -
Эта тема принадлежит разделу:
Предмет и методы генетики
Виды наследственности.. клетка как генетическая.. митоз..
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ:
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Все темы данного раздела:
Методы генетики
Специфические методы генетики.
1. Гибридологический метод (открытый Менделем). Основные черты метода:
а). Мендель учитывал не весь многообразный комплекс признаков у родите
Предмет генетики
Наследственность - это воспроизведение жизни (Н. П. Дубинин). Изменчивость - это возникновение различии между организмами по ряду признаков и свойств.
Наследственность, изменчивос
Виды изменчивости
Выделяют следующие виды изменчивости: мутационную, ком-бинативную, коррелятивную и модификационную.
Мутационная изменчивость. Мутация - стойкое изменение в структуре ДНК и кариотипе
Роль ядра и других органелл в прередаче, сохранения и реализации наследственной информации
Организм животных состоит из миллиардов клеток, и все они ведут начало от двух половых клеток (гамет) - отцовского спер-мия и материнской яйцеклетки. При слиянии гамет - о
Роль ядра и цитоплазмы в наследственности
Ядро - основной компонент клетки, несущий генетическую информацию. Оно может находиться в двух состояниях: покоя - интерфазы и деления - митоза или мейоза. Интерфазное ядро представляет собой кругл
Морфологическое строение и химический состав хромосом
При микроскопическом анализе хромосом прежде всего видны различия их по форме и величине. Строение каждой хромосомы сугубо индивидуальное. Можно заметить также, что хромосомы обладают общими морфол
Кариотип и его видовые ообенности
Анализ хромосом в клетках животных и растений разных видов позволил выявить ряд общих закономерностей, имеющих важное значение при изучении явлений наследственности и изменчивости. Установлено, чт
Роль генотипа и условий среды в формировании фенотипа
Фенотип каждого организма формируется под влиянием генотипа и условий среды. Генотип определяет норму реакции организма - границы изменчивости выражения признака под влиянием и
Особенности гаметогенеза самцов и самок
Процесс развития половых клеток носит название гаметогене-за. У самцов этот процесс называется спермиогенезом, а у самок - овогенезом (рис. б). Половые клетки в развитии после
Полиплоидия и ее значения
Полиплоидия (тут- «полп» -^* много, «плоид» -*- складывать) увеличение числа хромосом, кратное (эуплоидия) и некратное
(анэуплоидия) гаплойдному набору: 3н - триплоид 4н тетраплоид.
Паталогии мейоза и митоза и ее значения
Патология мейоза.Основная патология мейоза - нерасхождение хромосом. Оно может быть первичным, вторичным и третичным. Первичное нерасхождение возникает у особей с нормальным кар
Оплодотворение
Оплодотворение- процесс слияния яйцеклетки со сперматозоидом. Яйцеклетка^- женская гамета (половая клетка) -*- ужи-вотных образуется в ядониках. Она формируется в результате оогенеза и соде
Особенности гибридологического метода менделя
Сущность гибридологического метода заключается в следующем:
1) для скрещивания выбирают родительские формы, четко
различающиеся по одной, двум или трем парам контрастных,
Закон единообразия гибридов первого поколения
Мендель избрал для анализа семь пар четко различающихся признаков: форма семян - круглая или морщинистая, окраска семядолей - желтая или зеленая, окраска цветков - пурпурная или белая, рост растени
Закон расщепления
В следующем опыте путем скрещивания между собой гибридов первого поколения Мендель получил второе поколение, в котором наряду с доминирующими признаками проявились и рецессивные. Появление во втор
Аллели. Множественный аллелизм
Аллельными генами (аллелями) называют гены, расположенные в одинаковых точках (локусах) парных гомологичных хромосом. Аллели оказывают влияние на развитие одного и того же признака организм
Анализирующее скрещивание. Правило чистоты гамет
Мендель провел опыт по скрещиванию гибридов первого поколения с растениями гороха исходных родительских сортов. Скрещивание гибридов первого поколения (Аа) с особями, сходными по генотипу
Полигибридное скрещивание
Положение о независимом наследовании разных пар аллелей и признаков было подтверждено Менделем при изучении наследования трёх пар признаков у гороха. Он скрещивал сорт растения с круглыми семенам
Плейотропное действие генов
Плейотропия.Это влияние одного гена на развитие двух и более признаков (множественное действие гена). Так, Д. К. Беляев и А. И. Железнова установили, что у норок большинство мутац
Полимирия
Иногда на формирование признака влияют две или несколько пар неаллельных генов. Проявление признака в этом случае зависит от характера их взаимодействия в процессе развития организма. В первом по
Сцепленное наследование признаков
Мы познакомились с дигибридным скрещиванием и уяснили, что независимое комбинирование признаков объясняется тем, что расщепление одной пары аллельных генов, определяющих соответствующие признаки, п
Полное сцепление
Т. Морган скрещивал черных длиннокрылых самок с серыми с зачаточными крыльями самцами. У дрозофилы серая окраска тела доминирует над черной, длиннокрылость - над зачаточными крыльями. Обозначим ге
Неполное сцепление
В следующем опыте, так же как и в предыдущем, Морган скрещивал черных длиннокрылых самок с серыми зачаточно-крылыми самцами. В первом поколении получил все потомство серое длиннокрылое. Затем снова
Картирование хромосом
Сущность соматического кроссинговера заключается в том, что он осуществляется при митотическом делении соматических клеток главным образом эмбриональных тканей. Кроссинговер происходит меж
Карты хромосом
После того как была установлена связь генов с хромосомами и обнаружено, что частота кроссинговера всегда вполне определенная для каждой пары генов, расположенных в одной группе сцепления, встал во
Гомо и гетерогаметный пол
В ходе эволюции у большинства раздельнополых организмов сформировался механизм детерминации пола, обеспечивающий образование равного количества самцов и самок, что необходимо для
Наследование признаков, ограниченных полом
Наследование признаков, контролируемых генами, локализованными в аутосомах, но фенотипически проявляющихся исключительно или преимущественно у одного пола, - есть наследование, ограниченное п
Проблема регуляции пола
Проблема регуляции пола вытекает из необходимости увеличения продукции животноводства за счет преимущественного получения особей одного вида, дающих более высокий выход молока, мяса, шерсти, яиц и
Наследование признаков, сцепленных с полом
Причины более высокой смертности среди самцов млекопитающих можно объяснить исходя из особенностей наследования признаков, сцепленных с полом. Явление сцепленного с полом наследования впервые откр
Нуклеиновые кислоты, доказательства их роли в наслндственности
Генетическая информация реализуется в процессе биосинтеза белков. Все основные свойства живых существ определяются структурой и функцией белковых молекул. В последние 40 лет в ряде лаборатор
Химический состав и структура нуклеиновых кислот
Нуклеиновые кислоты впервые открыл И. Ф. Мишер в 1868 г. Он выделил из ядер клеток особое вещество кислотной природы и назвал его нуклеином. Впоследствии ему дали название «нуклеиновая кислота». Б
Днк рнк
Пуриновые основания Аденин, гуанин Аденин, гуанин
Пиримидиновые основания Цитозин, тимин Цитозин, урацил
Углеводный компонент Дезоксирибоза Рибоза
Об
Строение и типы рнк
Многочисленными исследованиями было установлено, что синтез белка в клетке происходит не в ядре, где находится ДНК, а в цитоплазме. Следовательно, сама ДНК не может служить матрицей для синтеза бе
Репликация ДНК
Репликация (удвоение) ДНК.ДНК находится в хромосомах, и репликация ее происходит перед каждым удвоением хромосом и делением клетки. Дж. Уотсон и Ф. Крик предложили схему уд
Генетический код и его свойства
Представление о том, что генетическая информация о структуре белковых молекул зашифрована в ДНК путем определенного расположения нуклеотидов, конкретизировал Ф. Крик в гипотезе последовательност
Синтез белка в клетке
В настоящее время можно считать установленным, что наследственность реализуется в процессе биосинтеза белка. Синтез ферментов и других белков, необходимых для жизнедеятельности и развития организм
Конъюгация
Конъюгация - перенос генетического материала от одной бактериальной клетки (донора) к другой (реципиенту) при их непосредственном контакте. Процесс конъюгации у бактерий обнаружили Дж. Ле
Трансдукция
Трансдукция - перенос генов из одной бактериальной клетки в другую при помощи бактериофага. Впервые это явление установили в 1952 г. Н. Зиндер и Дж. Ледерберг. Они проводили исследования на
Трансформация
Трансформация - поглощение изолированной ДНК бактерии донора клетками бактерии реципиента. Явление трансформации кратко освещено при изложении доказательств роли ДНК в наследственно
Мутационная изменчивость
В предыдущих главах отмечено одно из основных свойств кариотипа, ДНК и ее участков (генов) - сохранять постоянство внешнего и внутреннего строения. Морфофункциональная устойчивость генетического м
Хромосомные мутации
Изменения кариотипа могут быть количественными, структурными и одновременно теми и другими. Рассмотрим отдельные формы изменения хромосом (см. схему).
Числовые мутации кариотипа.
Генные мутации
По характеру действия генные мутации могут быть доминантными или рецессивными. Чаще мутантный ген обладает рецессивным эффектом. Нормальный аллель подавляет при этом действие измен
Регуляция синтеза ирнк и белка
Все клетки организма, как бы они ни были дифференцированы, как правило, тождественны по генотипу. В каждой клетке "имеются все хромосомы и весь набор свойственных данному организму генов. Однако к
Строение и размножение бактерий
Химический состав клеток бактерий в основном такой же, как и клеток высокоорганизованных организмов. Клетки бактерий окружены оболочкой, внутри которой находятся цитоплазма, ядерный аппарат, рибосо
Строение и размножение вирусов
Вирусы относятся к микроорганизмам, хотя резко отличаются от всех известных клеточных форм жизни. Частицы вирусов очень малы (от 20 до 450 нм). С помощью электронного микроскопа обнаружено, что он
Взаимодействие фага с бактериальной клеткой
К клеточной стенке бактерий фаги прикрепляются концевыми нитями отростков. Затем оболочка бактерии растворяется с помощью фермента лизоцима, белковый чехол хвостового отростка сокращается и через
Анеуплоидия
Числовые аномалии кариотипа (анеуплоидия). Числовые аномалии хромосом относят к вновь возникающим мутациям. Однако имеются исследования, которые показывают, что может быть семейная предрасположенно
Транслокации
Структурные мутации хромосом. Транслокации. Наибольшее количество исследований у крупного рогатого скота проведено по изучению частоты и влияния на плодовитость центрического слияния - тран
Генетические анамалии у с.х. животных
У сельскохозяйственных животных известны десятки аномалий, возникновение которых связано с рецессивными или доминантными мутациями генов. Эти аномалии встречаются в отдельных популяциях с разной
Простой аутосомный рецессивный тип наследования
Этим термином пользуются, когда аномалия обусловлена одним рецессивным геном, локализованным в аутосоме. Иногда употребляют термин моногенно-аутосомный тип наследования, что является синонимом. Ано
Аутосомный доминантный тип наследования
Признаки, обусловленные доминантными генами, как правило, проявляются в гетерозиготном состоянии. Возможный вариант скрещивания и характер расщепления следующие (табл. 40). При таком доминантном
Сцепленный с х-хромосомой тип наследования
В дополнение к главе «Генетика пола» отметим следующее. Гены, локализованные в Х-хромосоме, могут проявлять доминантный или рецессивный эффект. Возможные варианты скрещиваний и расщеплений при сц
Мультифакториальное наследование
Развитие исследований в генетике, в том числе и ветеринарной, привело к выводам о том, что далеко не при всех формах или даже случаях патология - простой менделирующий признак. Часто она обусловле
Практическое значение закона
ПОПУЛЯЦИЯ И «ЧИСТАЯ ЛИНИЯ»
По Н. В. Тимофееву-Ресовскому, популяция - это совокупность особей данного вида, в течение длительного времени (большого числа поколений
Структура свободно размножающейся популяции. Закон харди - вайнберга
Харда" и Вайнберг провели математический анализ распределения генов в больших популяциях, где нет отбора, мутаций и смешивания популяций. Они установили, что такая популяция находится в состоянии
Генная инженерия
Генная инженерия - раздел биотехнологии, связанный с целенаправленным конструированием in vitro новых комбинаций генетического материала, способного размножаться в клетке и синтезирова
Значение групп крови для практики
Контроль достоверности происхождения животных.Одна из главных областей практического применения групп крови - контроль происхождения животных. Такое их использование вызвано тем,
Значение биохимического полиморфизма
Биохимические полиморфные системы белков используются для следующих целей:
1) изучения причин и динамики генотипической изменчивости, составляющей основу эволюционной генетики;
2
Генетический груз популяций
В ходе длительной эволюции животных наряду с полезными мутациями, подхватываемыми отбором, в популяциях или породах накопился определенный спектр генных и хромосомных мутаций. Каждое поколение по
Методы получения трансгенных животных
Трансгеноз - экспериментальный перенос генов, выделенных из определенного генома или искусственно синтезированных, в другой геном. Животные, в геном которых интегрируют чужеродные гены, наз
Вам также будет интересно:

Наука как соц. институт
– сфера чел. деятельности, целью которой явл. изучение предметов и...

Потребность мировой экономики в минеральных ресурсах будет только расти. На этом фоне,...

Ген
-
структурная и функциональная
единица наследственности,
контролирующая развитие...

Взаимодействие
тел
«Ключом
ко всякой науке является вопросительный знак»
Оноре
де...

Имена советского происхождения - личные имена, бытующие в языках народов бывшего СССР,...

